biologi for Majors II
læringsresultater
- forstå tidligere og nye klassificeringssystemer for vira
fordi de fleste vira sandsynligvis udviklede sig fra forskellige forfædre, er de systematiske metoder, som forskere har brugt til at klassificere prokaryote og eukaryote celler, ikke særlig nyttige. Hvis vira repræsenterer” rester ” af forskellige organismer, er selv genomisk eller proteinanalyse ikke nyttig. Hvorfor?, Fordi vira ikke har nogen fælles genomisk sekvens, som de alle deler. For eksempel er 16S rRNA-sekvensen, der er så nyttig til konstruktion af prokaryote fylogenier, ikke til nogen nytte for et væsen uden ribosomer! Biologer har tidligere brugt flere klassificeringssystemer. Vira blev oprindeligt grupperet efter delt morfologi. Senere blev grupper af vira klassificeret efter den type nukleinsyre, de indeholdt, DNA eller RNA, og om deres nukleinsyre var enkelt – eller dobbeltstrenget. Imidlertid grupperede disse tidligere klassificeringsmetoder vira forskelligt, fordi de var baseret på forskellige sæt tegn på virussen. Den mest almindeligt anvendte klassificeringsmetode i dag kaldes Baltimore-klassificeringsskemaet og er baseret på, hvordan messenger RNA (mRNA) genereres i hver bestemt type virus.
tidligere klassificeringssystemer
vira indeholder kun få elementer, hvormed de kan klassificeres: det virale genom, typen af kapsid og konvolutstrukturen for de indhyllede vira. Alle disse elementer er tidligere blevet brugt til viral klassificering (tabel 1 og figur 1). Virale genomer kan variere i typen af genetisk materiale (DNA eller RNA) og dets organisation (enkelt – eller dobbeltstrenget, lineært eller cirkulært og segmenteret eller ikke-segmenteret). I nogle vira er yderligere proteiner, der er nødvendige til replikation, forbundet direkte med genomet eller indeholdt i det virale kapsid.
| tabel 1. Virusklassificering efter Genomstruktur og kerne | |
|---|---|
| Kerneklassifikationer | eksempler |
| RNA | rabiesvirus, retrovirus |
| DNA | herpesvirus, koppevirus |
| enkeltstrenget | rabiesvirus, retrovirus |
| dobbeltstrenget | herpesvirus, koppevirus |
| lineær | rabiesvirus, retrovirus, herpesvirus, koppevirus |
| cirkulære | papillomavirus, mange bakteriofager |
| ikke-segmenteret: genom består af et enkelt segment af genetisk materiale | parainfluensvirus |
| segmenteret: genomet er opdelt i flere segmenter | |
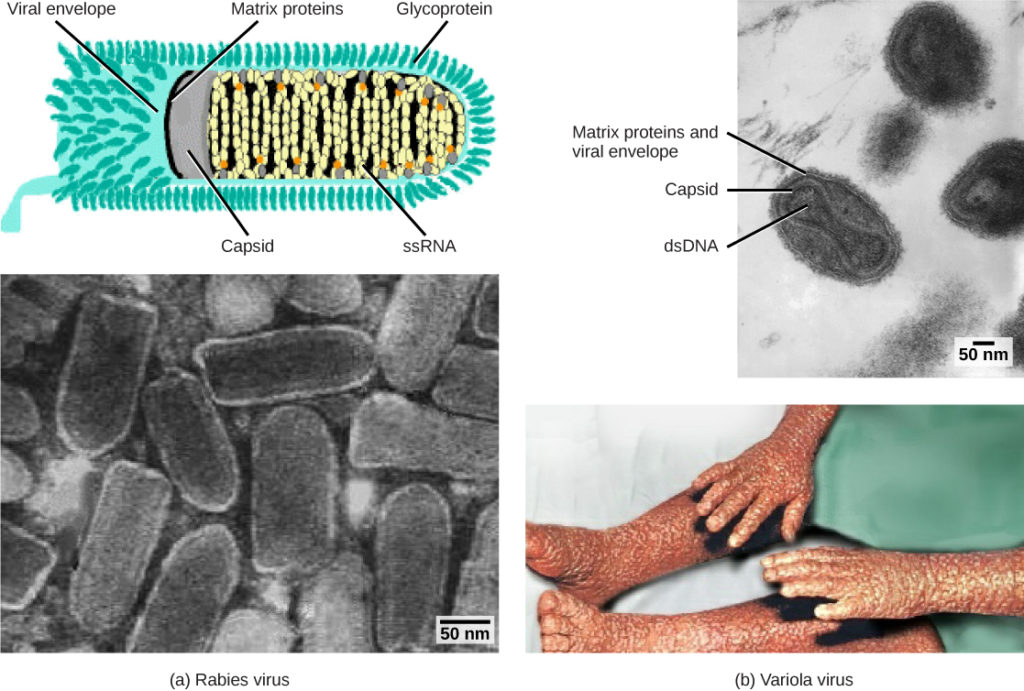
Figur 1. Vira klassificeres ud fra deres kernegenetiske materiale og kapsiddesign. (a) rabiesvirus har en enkeltstrenget RNA (ssRNA) kerne og en indkapslet spiralformet kapsid, mens (b) variola-virus, det forårsagende middel til kopper, har en dobbeltstrenget DNA (dsDNA) kerne og en kompleks kapsid. (kredit “rabies diagram”: ændring af arbejde af CDC; “rabies micrograph”: ændring af arbejde af Dr. Fred Murphy, CDC; kredit “lille koppemikrograf”: ændring af arbejde af Dr. Fred Murphy, Sylvia Hvidfield, CDC; kredit “koppefoto”: ændring af arbejde af CDC; skala-bar data fra Matt Russell)
virus kan også klassificeres ved udformningen af deres capsider (tabel 2 og figur 2). Capsider er klassificeret som nøgen icosahedral, indhyllet icosahedral, indhyllet spiralformet, nøgen spiralformet og kompleks. Typen af genetisk materiale (DNA eller RNA) og dets struktur (enkelt – eller dobbeltstrenget, lineært eller cirkulært og segmenteret eller ikke-segmenteret) bruges til at klassificere viruskernestrukturer (tabel 2).
| tabel 2. Virusklassificering efter Kapsidstruktur | |
|---|---|
| Capsid klassifikation | eksempler |
| nøgen icosahedral | Hepatitis A-virus, poliovirus |
| indkapslet icosahedral | Epstein-Barr virus, herpes virus, rubella virus, gul feber virus, HIV-1 |
| indkapslet spiralformet | virus af fåresyge, mæslingevirus, rabiesvirus |
| nøgen spiralformet | tobak mosaik virus |
| kompleks med mange proteiner; nogle har kombinationer af icosahedral og spiralformede kapsidstrukturer | herpesvirus, koppevirus, hepatitis B-virus, T4 bakteriofag |

figur 2. Transmissionselektronmikrografer af forskellige vira viser deres strukturer. Capsid af (A) poliovirus er nøgen icosahedral; (b) Epstein-Barr virus capsid er indhyllet icosahedral; (c) fåresyge virus capsid er en indhyllet spiral; (d) tobak mosaik virus capsid er nøgen spiralformet; og (e) herpesvirus capsid er kompleks. (kredit a: ændring af arbejde af Dr. Fred Murphy, Sylvia Hvidfield; kredit b: ændring af arbejde af Lisa Gross; kredit c: ændring af arbejde af Dr. F. A. Murphy, CDC; kredit d: ændring af arbejde af USDA ARS; kredit e: ændring af arbejde af Linda Stannard, Institut for Medicinsk Mikrobiologi, University of Kapstaden, Sydafrika, NASA; skala-bar data fra Matt Russell)
Baltimore Classification
det mest anvendte system for virusklassificering blev udviklet af Nobelprisvindende biolog David Baltimore i begyndelsen af 1970 ‘ erne. ud over forskellene i morfologi og genetik nævnt ovenfor grupperer Baltimore-klassificeringsordningen vira efter, hvordan mRNA produceres under virusets replikative cyklus.
gruppe i-vira indeholder dobbeltstrenget DNA (dsDNA) som deres genom. Deres mRNA produceres ved transkription på samme måde som med cellulært DNA.
gruppe II-vira har enkeltstrenget DNA (ssDNA) som deres genom. De konverterer deres enkeltstrengede genomer til et dsDNA-mellemprodukt, før transkription til mRNA kan forekomme.
Gruppe III-vira bruger dsRNA som deres genom. Strengene adskilles, og en af dem bruges som en skabelon til generering af mRNA ved anvendelse af den RNA-afhængige RNA-polymerase kodet af virussen.
gruppe IV-vira har ssRNA som deres genom med en positiv polaritet. Positiv polaritet betyder, at det genomiske RNA kan tjene direkte som mRNA. Mellemprodukter af dsRNA, kaldet replikative mellemprodukter, fremstilles i processen med at kopiere det genomiske RNA. Flere RNA-strenge i fuld længde med negativ polaritet (gratis til det positive-strandede genomiske RNA) dannes ud fra disse mellemprodukter, som derefter kan tjene som skabeloner til produktion af RNA med positiv polaritet, inklusive både genomisk RNA i fuld længde og kortere virale mRNA ‘ er.
gruppe V-vira indeholder ssrna-genomer med en negativ polaritet, hvilket betyder, at deres sekvens er komplementær til mRNA. Som med gruppe IV-vira bruges dsRNA-mellemprodukter til at lave kopier af genomet og producere mRNA. I dette tilfælde kan det negativt strandede genom omdannes direkte til mRNA. Derudover er positive RNA-tråde i fuld længde Lavet til at tjene som skabeloner til produktion af det negativt strandede genom.
gruppe vi-vira har diploide (to kopier) ssrna-genomer, der skal konverteres ved hjælp af revers transkriptase til dsDNA; dsDNA transporteres derefter til kernen i værtscellen og indsættes i værtsgenomet. Derefter kan mRNA produceres ved transkription af det virale DNA, der blev integreret i værtsgenomet.
gruppe VII-vira har partielle dsDNA-genomer og fremstiller ssrna-mellemprodukter, der fungerer som mRNA, men omdannes også tilbage til dsDNA-genomer ved revers transkriptase, der er nødvendig for genomreplikation. Karakteristika for hver gruppe i Baltimore-klassificeringen er opsummeret i tabel 3 med eksempler på hver gruppe.
| tabel 3. Baltimore klassifikation | |||
|---|---|---|---|
| gruppe | egenskaber | mRNA-produktionstilstand | eksempel |
| I | dobbeltstrenget DNA | mRNA transkriberes direkte fra DNA-skabelonen | herpesvirus) |
| II | enkeltstrenget DNA | DNA omdannes til dobbeltstrenget form, før RNA transkriberes | hundeparvovirus (parvovirus) |
| III | dobbeltstrenget RNA | mRNA transkriberes fra RNA-genomet | barndom gastroenteritis (rotavirus) |
| IV | enkeltstrenget RNA (+) | genom fungerer som mRNA | almindelig forkølelse (pircornavirus) |
| V | enkeltstrenget RNA ( – ) | mRNA transkriberes fra RNA-genomet | Rabies (rhabdovirus) |
| VI | enkeltstrengede RNA-vira med revers transkriptase | revers transkriptase fremstiller DNA fra RNA-genomet; DNA inkorporeres derefter i værtsgenomet; mRNA transkriberes fra det inkorporerede DNA | humant immundefektvirus (HIV) |
| VII | dobbeltstrengede DNA-vira med revers transkriptase | det virale genom er dobbeltstrenget DNA, men viralt DNA replikeres gennem et RNA-mellemprodukt; RNA kan tjene direkte som mRNA eller som en skabelon til fremstilling af mRNA | Hepatitis B-virus (hepadnavirus) |