Buněčná biologie 06: cytoskelet Část II: Tubulin
toto jsou poznámky z přednášky 6 kurzu buněčné biologie Harvard Extension.
minulý týden jsme pokryli mikrofilamenty, které jsou vyrobeny z aktinu. Tento týden: mikrotubuly vyrobené z tubulinu. Proč se ptáte, potřebuje cytoskeleton dva samostatné systémy? Zvažte, že mikrofilamenty jsou jen řetězy jednotlivých podjednotek, zatímco mikrotubuly jsou doslova trubky (jak uvidíme brzy) – duté válce se stěnami z řetězců vyrobených z dimerů. Tyto různé struktury znamenají různé schopnosti: mikrovlákny mohou snadněji tvořit spontánně, a může větev (s pomocí Arp2/3 a jiné komplexy mluvili minule) za účelem vytvoření různých síťových tvarů a připojení různých částí buňky. Mikrotubuly více spoléhají na nukleace v zájmu formě, a jsou v podstatě mluvil sítě spojující centrosome na okraj buněk. V praxi mikrovlákny jsou bohaté na buňky kůry a silně podílí kontraktilní pohyby a pohyblivosti buněk, zatímco mikrotubuly jsou nejvíce podílí na organizaci organel a slouží jako dráhy pro anterográdní a retrográdní transport.
Mikrotubuly
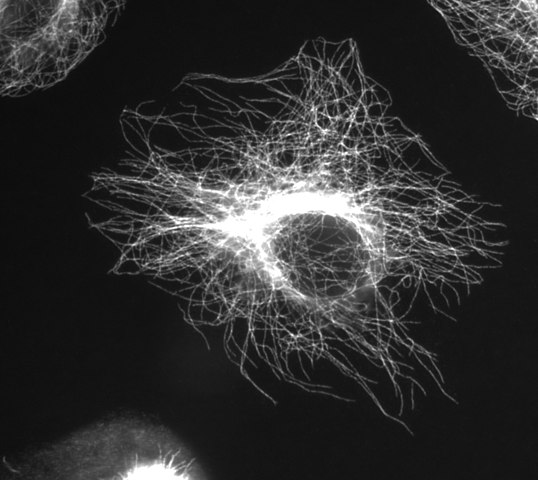
(Obraz díky Wikimedia Commons user Jeffrey81)
kromě Wikipedie, předchozí vydání Lodish a Cooper buněčné biologie učebnic dostupných na NCBI jsou také výborné reference.
trubková struktura mikrotubulů je robustnější než mikrovlákna, což umožňuje mnoho těžkých tahů a tlačení. I když jsou mikrotubuly robustní, jsou závislé na teplotě-depolymerují, pokud jsou ochlazeny na 4°C, a znovu se repolymerují, pokud jsou zahřáté na 37°C za předpokladu, že je k dispozici GTP.
základním stavebním kamenem mikrotubulů je dimer α-tubuilinu/β-tubulinu (kódovaný geny TUBA_ a geny TUBB_). Jednotlivé tubulinové proteiny váží ~55kDa, takže při ~110Da / aminokyselině je to řádově 500 aminokyselin. Α – i β-tubulin váží GTP, ale α se na něm drží navždy, kdežto v β může být vyměněn nebo hydrolyzován na GDP a poté znovu vyměněn za nový GTP.
Prameny po sobě jdoucích α/β dimery tvoří protofilament a 13 protofilament uspořádány side-by-side do válce, aby mikroskopický. Prostor mezi protofilamenty se nazývá „šev“. Celý mikrotubul má průměr ~25 nm. Jeho strukturu (tvořenou dimery a protofilamenty) můžete vidět ve vnitřním životě buněčného segmentu kolem mikrotubulů:
v Rámci každého dimeru, β podjednotka je (+) konec, oblíbené pro polymerace a α je (-) konec, oblíbené pro depolymerací. Mikrotubuly se tvoří v podstatě ve stejných třech krocích jako mikrofilamenty-nukleace – prodloužení a ustálený stav. Na rozdíl od mikrofilamentů se však nukleují samy o sobě snadno, takže nukleace vyžaduje centra organizující mikrotubuly (Mtoc). Nerozdělující se buňky mají každý pouze jeden MTOC, nazývaný centrosom. (To je také znázorněno na videu výše). Nachází se v blízkosti jádra, centrosome je rozbočovač pro radiální configuation mikrotubulů s (-) konce poukázal v a (+) koncích směřující k periferii buňky.
centrosom se skládá ze 2 válců nazývaných centrioly. Každý centriole se skládá z jednotlivých 9 sad 3 bočně srostlé mikrotubuly, obklopené amorfní ‚pericentriolar materiál bohatý na věci, které podporují nukleace – zejména γ-tubulin ring komplexy (gama-tubulin je kódován podle TUBG_ geny). γ-tubulin je považován za „dělenou podložku“:

model je, že roztřepené konečky umožňují γ-tubulin se vázat na α-tubulin – tj. (-) konci k-být-tvoří mikroskopický – zajištění nukleace osiva pro mikroskopický tvořit.
mikrotubuly mohou být vytvořeny in vitro. Dynamika závisí hlavně na kritických koncentracích. (- ) Konec je méně nakloněn k polymeraci než ( + ) konec, takže má vyšší kritickou koncentraci. Pokud je skutečná koncentrace mezi kritickými koncentracemi obou konců, dochází ke šlapání.
když se mikrotubule náhle začne disociovat, nazývá se to „katastrofa“. Katastrofy mají zajímavou energetickou dynamiku. Připomeňme si dříve, že beta-tubulin, což je (+) konec, kde dochází k prodloužení, může být vázán buď na GTP nebo na HDP. Je stabilnější součástí jeho mikrotubulů, když je vázán na GTP; tubulin vázaný na HDP je schopen disociovat. Mikrotubuly tvoří především přidáním GTP-vázané β-tubulin na (+) konci, ale poté, co byl přidán beta-tubulinu později rozpouštějí jejich GTP, takže je HDP-vázané. Takže tam je druh “ tip “ mikrotubulové, že je GTP-vázané, zatímco beta-tubulinu hlouběji mikrotubulové, přidáno delší lety, jsou HDP-vázané. Pokud hranici GTP hydrolýza úlovky až do špičky, nebo když se něco stane na sever mikrotubulové, pak méně stabilní, HDP vazbou beta-tubulinu se stává vystaveny, a protofilament začne loupat pryč, jako „beraní rohy“. Pokud k tomu dojde, mikrotubule se rozebere, dokud nenarazí na „ostrov“ beta-tubulinu vázaného na GTP někde dále po slámě. Katastrofa může být „zachránil“ přidáním nových GTP-vázané tubulinových dimerů, že bude čepice konce a stabilizovat protofilament, umožňující mikroskopický re-forma. Můžete vidět v tomto videu, že depolymerizace mikrotubulů může být mnohem rychlejší, než polymerace:
Zde jsou některé sloučeniny užitečné pro studium mikrotubulů. Kolchicin, lék proti dnu, váže volné alfa-beta dimery, snižuje jejich přísun pro tvorbu mikrotubulů a tím podporuje depolymeraci. Nocodazol také interferuje s tvorbou nových mikrotubulů a protože tvorba nových mikrotubulů je důležitá pro mitózu, nocodazol je protinádorový lék proti rakovině. Naopak paklitaxel (taxol), další protirakovinné léčivo, působí stabilizací mikrotubulů, protože rozpad stávajících mikrotubulů je také důležitý pro mitózu.
proteiny spojené s mikrotubulemi (mapy) mají různé role. Pozoruhodné jsou MAP4 (v neuronálních buňkách), MAP2 (v neuronech) a Tau (v neuronech; kódováno genem MAPT). Všechny tyto mají společné to, že působí na stabilizaci mikrotubulů, posouvají kinetiku ve prospěch růstu mikrotubulů a proti katastrofě. Každá obsahuje 18 aminokyselinový úsek s pozitivně nabitými aminokyselinami, které se vážou na negativní část mikrotubulů. Mohou působit na Svazování více mikrotubulů dohromady , přičemž MAP2 drží mikrotubuly ve větší vzdálenosti díky delšímu rameni (ve srovnání s Tau).
mapy jsou regulovány fosforylací: MARK protein kinázy kovalentně vážou fosfátové skupiny na S, T nebo Y aminokyseliny v proteinech MAP, což snižuje jejich schopnost vázat mikrotubuly. (Cyklin dependentní kinázy také regulují mapy během mitózy). Předpokládá se, že tyto mapy slouží k určení fyzické struktury buněk. V neuronech se MAP2 nachází v dendritech a Tau je většinou v axonu. Mutace v genu MAPT, který kóduje Tau, způsobují frontotemporální demenci (FTD). Hyperfosforylovaný Tau se nachází (i když nevíme proč) v mozcích pacientů s Alzheimerovou chorobou. Myší modely obou těchto onemocnění ukazují, axonální degenerace , v souladu s Tau není schopen dělat svou práci na stabilizaci mikrotubulů v axonu.
další třída proteinů, pojmenovaná +Tipy pro jejich vazbu na (+) konec mikrotubulů, může chránit před katastrofou. Zdá se, že rozšiřují spravedlivé cesty dolů do mikrotubulů. Toto video ukazuje, že EB-1 je tlačen ven, protože konce mikrotubulů rostou:
jiné proteiny vázající se na konec podporují spíše katastrofu než stabilitu. Kinesin-13 působí na zakřivení konce protofilamentů a snižuje prahovou hodnotu pro katastrofu. Stathmin (STMN1 gen; aka Op18, kde Op je zkratka pro onkoproteinu) se váže na tubulinových dimerů v protofilament, podpora obou bezprostřední katastrofu a možná také GTP hydrolýzy, které tím, že odstraní „ostrovy“ GTP-vázané beta-tubulin by představovalo více dlouhodobé investice ve prospěch katastrofa. Stathmin je regulován fosforylací. Katanin (po japonském meči katana) doslova odděluje mikrotubuly.
motorické proteiny mikrotubulů jsou do značné míry podobné motorickým proteinům mikrofilamentů. Přicházejí ve dvou rodinách: kinesin, z nichž většina pohybovat anterográdní tj. směrem k (+) konci, a dyneins, z nichž většina pohybovat retrográdní tj. směrem k (-) konci. „Chodí“ takto:
Kinesin a dynein se podílejí na pohybu organel, endocytóze a exocytóze a segregaci chromozomů během meiózy & mitózy.
Kinesin-1, nejlépe studovaný z kinesinů, je „konvenční“ kinesin v tom, že působí jako tetramer složený ze dvou jednotek těžkého řetězce (např. KIF1A), které tvoří „hlavy“ domén, které rozpouštějí ATP a vázat mikrotubuly a dva lehké řetězce jednotek (KLC geny např. KLC1), které tvoří „ocas“, který se váže náklad, v tomto případě váčků. Linker doména(který protein je tato část?) umožňuje dimerizaci dvou těžkých řetězů.
zde je video z toho v akci. Všimněte si, že video volá dimer místo tetrameru; myslím, že je to proto, že zvažují pouze hlavy a nediskutují o ocasech.
některé další kineziny se trochu liší:
- Kinesin-2, který také dělá váček & organely, transport, je heterotrimer dvě odlišné (i když související) těžkých řetězců a další polypeptid používá k regulaci jeho nákladu.
- Kinesin-5 má hlavy na obou koncích místo hlavy a ocasu, takže to chodí v protilehlých směrech na dvou různých mikrotubuly, tahem k sobě. Tomu se říká „bipolární hnutí“.
- Kinesin-14 je jediný známý kinesin, který se pohybuje směrem k ( – ) konci a podílí se na mitóze.
zásady „chůze“ pohybu jsou skoro stejné pro všechny kinesin, i když, a jsou to, co je líčen v posledním videu výše. Nejsou-li vázány na mikrotubulu, jsou „hlavy“ vázány na ADP. Jedna hlava se stane, že narazí na mikrotubulu a naváže se na ni, uvolní její ADP, což umožní ATP, aby ji nahradil. ATP vazba indukuje konformační změnu, která táhne na linker, kyvné jiné, zaostávající hlavu dopředu, do vedoucí pozice, kde se váže na mikroskopický. Původní hlava pak hydrolyzuje ATP, který uvolňuje fosfát (což je zkráceně Pi ve videu) a poskytuje energii pro jeden energicky do kopce krokem v tomto procesu, který je oprostit se od mikrotubulové.
Myosin (motorický protein, který chodí na mikrofilamenty) a kinesin mají velmi podobné struktury, ale žádná podobnost sekvence aminokyselin. Proto se předpokládá, že nejsou paralogové, ale spíše příklad konvergentní evoluce.
lidé mluví hodně o kinesins částečně proto, že nerozumíme dyneins docela dobře. Dyneiny jsou obrovské proteiny, vyrobené ze 2 velkých, 2 středních a 2 malých podjednotek. Jejich pouhá velikost je znesnadnila izolovat a charakterizovat a jejich fungování není dobře pochopeno. Víme, že dynaktinový komplex (multiproteinový komplex; samotné dynaktiny jsou geny DTCN_) je zapojen jako „adaptér“, který spojuje dynein s nákladem.
toto video shrnuje celý cytoskelet a lana ve většině materiálů z minulého týdne a této přednášky:
PrP
Jak je uvedeno v sekreční dráhy poznámky, PrP je GPI-ukotveny k membráně a prochází endocytóza velmi pravidelně, vytváří endocytic váčky s PrP v nich. Encalada 2011 zjistí, že Kinesin-1C a DHC1 (dynein heavy chain 1)jsou zodpovědné za přepravu těchto PRP vezikul anterográdní a retrográdní. Když Encalada vyrazil, srazil nebo inhiboval Kinesin-1C, anterográdní a retrográdní pohyb byly sníženy a podobně, když byl narušen DHC1. Zdá se tedy, že tyto dva komplexy, i když se pohybují v opačných směrech, se navzájem aktivují. A zajímavé je, že narušení těchto motorických proteinů nezabránilo tomu, aby se vezikuly PrP spojily s motory – prostě se nepohybovaly tak rychle nebo tak často. Papír má řadu dalších buněčné biologie závěry o povaze, jak váčků aktivovat motorické proteiny a jak směr dopravy je určena.