Hranic, v Mikrobiologii
Úvod
Astroviridae rodina obsahuje neobalené, pozitivní smysl, single-stranded RNA viry do dvou rodů, Mamastrovirus a Avastrovirus, které infikují savce a ptáky, resp. V současné době, Mezinárodní Výbor pro Taxonomii Virů (Mezinárodní Výbor pro Taxonomii Virů , 2018) uznává, 19 druhů, a to Mamastrovirus-1 až -19, ve Mamastrovirus rodu; nicméně, tam jsou četné kmeny čeká klasifikace, z nichž některé jsou předběžně považovat za nový druh (Donato a Vijaykrishna, 2017).
od roku 2010 je několik astrovirů stále více uznáváno jako neuroinvazivní u různých druhů savců, včetně lidí (Quan et al ., 2010; Naccache a kol., 2015), norek (Blomström et al., 2010), dobytek (Li et al., 2013), Ovce (Pfaff et al., 2017) a prasata (Boros et al., 2017). Po počátečním rozpoznání bovinní encefalitidy spojené s astrovirem u skotu Spojených států (Li et al ., 2013), retrospektivní studie případů sporadické skotu encefalitidy neurčené etiologie ze Švýcarska ukázal, že to neuroinvasive astrovirus šel nepozorovaně po celá desetiletí (Selimovic-Hamza et al., 2016). I když epidemiologie a přenos tras z těchto astroviruses jsou neznámé, cross-druh přenosu bylo navrženo na základě vysoké úrovni identity (>98%), sdílené mezi skotu a ovcí neuroinvasive astroviruses na nukleotidové a aminokyselinové úrovni (Boujon et al., 2017).
bovinní astroviry (BoAstVs), pojmenované BoAstV-NeuroS1 (Li et al., 2013) a BoAstV-CH13 (Bouzalas et al., 2014), byly původně nalezeny v mozku skotu s nehnisavou encefalitidou ve Spojených státech a Švýcarsku. Přes rozdílnou nomenklaturu představují oba viry stejný genotypový druh (Bouzalas et al., 2016; Selimovic-Hamza a kol., 2017a), která stále čeká na oficiální klasifikaci ICTV. V roce 2015 byl v mozku krav s encefalitidou ve Švýcarsku identifikován dříve neznámý kmen BoAstV, pojmenovaný BoAstV-CH15. Kompletní genom fylogenetické srovnání odhalilo bližší vztah BoAstV-CH15 s ovcí astrovirus (OvAstV) než s BoAstV-CH13 (Seuberlich et al., 2016). Koinfekce s BoAstV-CH13 a BoAstV-CH15 byla také dokumentována v jednom případě (Seuberlich et al., 2016). Ve stejném roce v Německu, Schlottau et al. (2016) uvádí, román astrovirus, a to BoAstV-BH89/14, v krávě, encefalitida, která byla nejvíce úzce souvisí s OvAstV a BoAstV-CH15. Následně byl v roce 2017 identifikován BoAstV-CH13/NeuroS1 v případech bovinní encefalitidy ve východní a západní Kanadě (Spinato et al ., 2017; Selimovic-Hamza a kol., 2017b). V roce 2018, román neuroinvasive BoAstV úzce souvisí s Severní Ameriky a Evropské BoAstV-NeuroS1/BoAstV-CH13, byl označen za vola s non-hnisavé encefalomyelitida v Japonsku a vzniku intra-genotypové rekombinace mezi Severní Ameriky a Evropské kmeny bylo navrhl, (Hirashima et al., 2018).
Zatímco případy astrovirus-spojené encefalitidy bylo hlášeno v Severní Americe, Evropě a Asii, jejich přítomnost nebyla nikdy zdokumentována na Jižní polokouli. Zde popisujeme případ astrovirus-spojené encefalitidy v dobytek v Uruguayi, která rozšiřuje geografické rozložení a genetické rozmanitosti neuroinvasive astroviruses a poskytnout phylogeographic důkazy, které naznačují, že tento virus byl představen do Ameriky z Evropy a později se rozšířil do Asie.
Materiály a Metody
Historie a Signalment
V červnu 2018, 22-month-old Holstein řídit ve skupině 37 voli v ∼300 ha zemědělské v Colonia, Uruguay, vyvinutý progresivní neurologické zpívá, včetně neobvyklé chování, bezcílné chození, kroužení, ataxie, opakující se a nekoordinované pohyby jazyka a poloze. Stádo se paslo na roční ovsené pastvině a bylo doplněno kukuřičnou siláží. Předpokládaná klinická diagnóza mozkové listeriózy veterinárním lékařem vedla k léčbě penicilinem a streptomycinem, zvíře však spontánně zemřelo po klinickém průběhu trvajícím 3 dny.
Patologické Vyšetření, in situ Hybridizace (ISH) a Imunohistochemie (IHC)
vedoucí řídit byl vyjmut z jatečně upraveného těla a předložen INIA je Veterinární Diagnostické Laboratoři (Zdraví Zvířat Platforma) pro diagnostické práce-up. Polovinu mozku, krátký úsek proximální krční míchy (C1), trigeminální ganglia a kořene trojklanného nervu, slinné žlázy, retrofaryngeální lymfatických uzlin, hltanu, jícnu, jazyka a kosterního svalstva, byli ponoření-opraveno v 10% neutrálním pufrovaném formalínu po dobu 48-72 h. Tkáně byly pravidelně zpracovávány pro histologie, vložené do parafínu, mikrotom-dělené na 4-5 µm a obarveny hematoxylinem a eosinem (H&E) a Gram skvrn.
Chromogenní ISH byla provedena ručně na 5 µm části formalin-fixed, paraffin-embedded (FFPE) mozkového kmene, mozečku a mozku na Superfrost Plus skluzavky (Thermo Fisher Scientific, Pittsburgh, PA, Spojené Státy) pomocí RNAscope 2.5 Red assay kit (Cat #322360, Advanced Cell Diagnostics, Hayward, CA, Spojené Státy) a BoAstV sonda Kočka. #406921. Sonda se skládá z 20 párů zaměřených na oblast 5232-6180 viru (GenBank KF233994. 1). Každá 5 µm část tkáně byla před hybridizací sondy předběžně ošetřena teplem a proteázou po dobu 2 hodin při 40°C a zpracována podle doporučení výrobců. Negativní kontroly používané pro validaci signálu zahrnuty nesouvisející (GC-obsah uzavřeno) sonda běh na sériové řezy a sondování tkáně z neinfikovaných zvířat. Diapozitivy byly kontrastovány s hematoxylinem a namontovány s EcoMount (Biocare Medical, Concord, CA, Spojené státy americké).
kromě toho byl IHC proveden v sekcích FFPE mozkového kmene, mozku a mozečku, jak bylo dříve popsáno, pro identifikaci viru Západonilského (WNV, Flavivirus) (Palmieri et al ., 2011), virus vztekliny (Lyssavirus) (Stein et al., 2010) a Chlamydia spp. (Giannitti et al., 2016) antigeny.
Molekulární Virologie
Nukleové kyseliny, extrakce byla provedena ze směsného vzorku zmrazené (-20°C) mozku pomocí MagMAX Nukleové Kyseliny Izolace Kit® (Thermo Fisher Scientific). Pro astrovirus detekce, reverzní transkripce (RT) byla provedena s RevertAid Reverzní Transkriptázy® (Thermo Fisher Scientific) a random hexamer primerů (Qiagen). PCR byla provedena z cDNA pomocí MangoMix® (Bioline) a primery, které zesilují 432-nukleotidů fragment astrovirus polymerázové gen (Tse et al., 2011). Produkt PCR byl vizualizován ve 2% agarózovém gelu, čištěn pomocí Purelink® Quick gel Extraction and PCR Purification Combo Kit (Invitrogen) a sekvenován v Macrogen Inc. (Soul, Jižní Korea). Pro astrovirus celého genomu amplifikace, Maxima H Minus Reverzní Transkriptázy (Thermo Fisher Scientific) a oligo(dT)18 pro pořízení z cDNA, a MangoMix® (Bioline) nebo Ranger DNA Polymerase (Bioline) s primery popsané Hirashima et al., 2018, byly použity. Produkty PCR byly vizualizovány v 1-2% agarózovém gelu, čištěny a sekvenovány, jak je uvedeno výše. Sestava sekvence byla provedena se Seqmanem (Lasergene 8, DNASTAR). Dvacet-šest kompletních genomových sekvencí neuroinvasive astrovirus ze skotu, ovcí, prasat, člověka a norek, a střevní skotu astrovirus k dispozici v GenBank byly staženy a zarovnány pomocí Clustal W v MEGA 7 software (Kumar et al., 2016). W-IQ-TREE1 (Trifinopoulos et al., 2016) byl použit k určení nejlepší-fit model posloupnost evoluce (SYM+I+G4) a vytvořit maximální-pravděpodobnost, že fylogenetický strom s téměř kompletní sekvence BoAstV zjištěny v tomto případě, a těch kompletních sekvencí stažených z GenBank pomocí bootstrap jako statistická metoda k posouzení clades robustnost. Podobnost plot byl proveden s SimPlot software (lol et al ., 1999). P-vzdálenosti na úrovni aminokyselin ORF2 byly odhadnuty pomocí softwaru MEGA 7 (Kumar et al ., 2016).
navíc byla provedena Bayesovská fylogeografická analýza s balíčkem BEAST v1.8.4 (Drummond et al., 2012), použití: kompletní kódování regionu BoAstV CH13/NeuroS1 linie, ORF1ab (non-strukturální geny), ORF2 (strukturní geny), ORF1a (proteázy) a částečné ORF1b (polymerázová genomické oblasti, pro které Kanadské kmeny byly k dispozici), všechny sekvence jsou k dispozici v GenBank (poslední přistoupení 18. dubna 2019), k určení evoluční rychlosti, věk/roky společné předky, a nejpravděpodobnější trasa virové oběhu země (Švýcarsko, Uruguay, usa, Kanada a Japonsko). Nedostatek rekombinace v datovém souboru byl stanoven pomocí Rekombinačního detekčního programu 4. Substituční model, který nejlépe vyhovuje každému zarovnání, byl stanoven pomocí softwaru MEGA 7 pomocí hodnot Bayesovského informačního kritéria (BIC) a časová struktura každé datové sady byla vyhodnocena pomocí TempEst (Rambaut et al ., 2016). X uvolněný molekulární hodiny s Bayesian Skyline analýzu byl vybrán Bayes Faktor mezi různými kombinacemi molekulární hodiny a splývající strom záznam použit. Země detekce byla použita jako rys. Na Markov chain Monte Carlo délka byla 100 milionů generací, zajištění konvergence analýzy vyhodnoceny v Tracer v1.6.0 a zadní pravděpodobnost byla použita k vyhodnocení clades. Maximální clade důvěryhodnost strom (MCCT) byla získána pomocí TreeAnnotator software od zvířete a zobrazil v FigTree v1.4.3.
nakonec byla DNA extrahovaná ze zmrazeného mozku zpracována pomocí PCR pro detekci bovinních herpesvirů 1 a 5 (BHV-1 a -5), jak bylo dříve popsáno (Ashbaugh et al., 1997).
Bakteriologie
Čerstvé vzorky mozku a mozkového kmene byly pravidelně zpracovávány pro aerobní bakteriální kultury v krvi a MacConkey lze testovat kolonie, a selektivní kultury pro Listeria monocytogenes (Al-Zoreky a Sandine, 1990).
Výsledky a Diskuse
klinických příznaků a epidemiologických zjištění v případě zde popsaných, byť non-specifické, byly podobné těm, které jsou popsány v jiných případech skotu astrovirus-spojené encefalitidy, který je obvykle popisován jako sporadické (Selimovic-Hamza et al., 2016), s různými neurologickými deficity (Deiss et al., 2017), s trváním klinických příznaků, které se obvykle pohybují od 1 dne do 3 týdnů (Schlottau et al ., 2016; Deiss et al., 2017; Spinato a kol., 2017; Hirashima a kol., 2018).
makroskopické vyšetření mozku, segmentu C1 míchy a dalších tkání hlavy neodhalilo významné hrubé anatomické léze. Histologicky, tam byl středně těžké až těžké, lymfocytární, histiocytární a plasmacytic meningoencephalomyelitis ovlivňující telencephala (včetně mozkové hemisféry a hippokampus), mozkový kmen, a jen zkoumal segmentu míchy. Léze byly převážně distribuovány v šedé hmotě a omezujících oblastech bílé hmoty. V postižených oblastech bylo perivaskulární přehýbání a lymfoplasmacytická a histiocytární zánět a neuronální nekrózy/neuronophagia s gliosis v přilehlé neuropil. Došlo k satelitóze postižených nekrotických neuronů (obrázky 1A-D). Léze byly mnohem méně časté a závažné v mozkovém parenchymu, i když došlo k multifokální středně cerebelární leptomeningitidě. Nebyly nalezeny žádné intralezionální bakterie S H&e a gramovými skvrnami. V ostatních vyšetřovaných tkáních nebyly zjištěny žádné významné histologické změny.

Obrázek 1. Histologické léze v mozkovém kmeni (A,B) a mozkové kůry (C,D) a detekce BoAstV RNA v mozkové kůře (E,G). Obrázky (A-D) jsou části mozku obarvené H&E; obrázky E a G jsou úseky mozkové kůry prokazující hybridizace pomocí chromogenního ISH pomocí BoAstV specifické sondy, kontrastním hematoxylinem; obraz F je sériový části mozkové kůry DapB sondy (negativní kontrola), kontrastním hematoxylinem. (A) perivenulární prostor je výrazně rozšířen zánětlivými buňkami (většinou lymfocyty a histiocyty), které také infiltrují sousední neuropil. (B) neuron v centru má hypereozinofilní perikarya a karyorrhexis (nekróza) a neuronální tělo je obklopen zvýšení počtu gliových (satellitosis) a zánětlivých buněk. C) leptomeninge je infiltrován lymfocyty a histiocyty. (D) oblasti mozkové kůry s více hypereozinofilní (nekrotické) neuronů a velké nádoby s perivaskulární lymfocytární manžety. (E) V sériové část mozkové kůry, bohaté intracytoplazmatická BoAstV RNA, značení je znázorněno silný, granulované červené chromogen depozice v cytoplazmě neuronů soma a neuronální rozšíření (E,G), že není přítomen hybridizace s použitím negativní kontroly sondy (F).
při histologickém vyšetření centrálního nervového systému bylo podezření na neuroinvazivní virovou infekci. Skot s encefalitidou je znepokojen, protože mnoho neuropatogenů přežvýkavců je zoonotických (Cantile a Youssef, 2016) ; diagnóza encefalitidy by tedy měla vést k rozsáhlému laboratornímu testování, aby bylo možné vyšetřit infekční agens, pokud je to možné. V případě zde popsaném, IHC pro WNV, virus vztekliny a Chlamydia spp. a PCR pro BHV-1 a -5 byly všechny negativní a z mozkové tkáně nebyly kultivovány žádné patogenní bakterie. Protože řídit < 2 roky staré a ne spongiformní změny byly pozorovány v mozkovém kmeni, bylo zvíře nejsou testovány na bovinní spongiformní encefalopatie (BSE), což je exotické nákazy z dospělého skotu, který nebyl nikdy hlášen v Uruguayi. BSE navíc není zánětlivá (Cantile a Youssef, 2016).
In situ hybridizace byla provedena pomocí sondy generovány z BoAstV-NeuroS1, a tam byl sondy hybridizace bohaté uvnitř, a omezena na cytoplazmy neuronů v mozkové hemisféry a hippokampus (Obr. 1E–G). V těchto oblastech se hybridizace sondy kolokalizovala s nekrotickými neurony a oblastmi gliózy, bez hybridizace sondy detekovatelné v gliových buňkách nebo zánětlivých buňkách perivaskulárních manžet. Žádné virové nukleové kyseliny byl detekován ISH v mozečku, který měl jen minimální zánětlivé léze v parenchymu ale mírné leptomeningitis, nebo mozkového kmene, včetně úseků s těžkým zánětem. To znamená, že, topograficky, detekce virových distribuce ISH byl omezen více, než encefalitida v sekcích zkoumal, které se příležitostně byly popsány v případech BoAstV-CH13/NeuroS1-spojené encefalitidy u skotu (Selimovic-Hamza et al., 2017a, b). Důvod pro tento občasný nedostatek virové RNA detekce v lesioned oblastech mozku může být detekční limit ISH, nebo clearance viru v zanícené oblasti mozku, o době smrti, jak již dříve navrhl (Selimovic-Hamza et al., 2017b). Jak se očekávalo, žádná hybridizace sondy nebyla detekována ISH v mozkové tkáni použité jako negativní kontrola.
Astrovirus byl detekován v mozku RT-PCR. Téměř kompletní sekvence genomu, analýza odhalila Mamastrovirus napětí v CH13/NeuroS1 clade, pojmenovali jsme BoAstV-Neuro-Uy, sekvence byla uložena v GenBank pod přistoupení číslo MK386569. Na fylogenetická analýza ukázala, blízkost s jinými neuroinvasive astroviruses ve Virginii/Human-Norek-Ovcí (VA/HMO) clade (Obrázek 2), který obsahuje nejznámější neuroinvasive astroviruses (Hirashima et al., 2018; Reuter a kol., 2018). Téměř úplná sekvence BoAstV-Neuro-Uy je 6427 bp na délku a má sekvenční identitu 94% s kmenem KagoshimaSR28-462. BoAstV-Neuro-Uy má podobné vlastnosti jako jiné kmeny rodu CH13/NeuroS1: 5’UTR oblasti 51 nt, ORF1a (proteázy) 861 aminokyselin (aa), ORF1b 523 aa (RNA-dependentní RNA polymeráza), a ORF2 758 aa (kapsidový protein). Bohužel, 3 ‚ UTR nemohl být sekvenován, ale předpokládá se, že je přítomen poly(a) ocas, protože oligo (dT)18 byl použit k získání cDNA. Kromě toho je přítomna heptamerická sekvence AAAAAAC, ribozomální signál posuvu snímků. P-vzdálenosti na úrovni aminokyselin ORF2 potvrdily přiřazení tohoto kmene k clade CH13/NeuroS1. P-vzdálenosti < 0.35 mezi BoAstV-Neuro-Uy a další členové tohoto kmene (Tabulka 1) podpora klasifikace tyto virové kmeny v rámci jednoho stejného druhu; Mamastrovirus-13 byl nedávno navržených jinými autory (Donato a Vijaykrishna, 2017; Hirashima et al., 2018), ačkoli definitivní přiřazení druhů ICTV čeká. Sonda použitá pro ISH, generovaná z BoAstV-NeuroS1, měla 92,7% sekvenční identitu s BoAstV-Neuro-Uy.
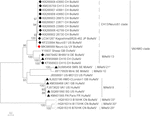
Obrázek 2. Fylogenetická analýza celovečerních nukleotidových sekvencí metodou maximální věrohodnosti. BoAstV-Neuro-Uy je označen červeným kosočtvercem. Další sekvence skotu neuroinvasive astroviruses jsou označeny černé kosočtverce, sekvence z non-skotu neuroinvasive astroviruses jsou označeny černými trojúhelníky, a sekvence z enterické skotu astroviruses jsou označeny bílými kruhy. Jsou zobrazeny druhy mamastrovirů; hvězdičky označují druhy, které ICTV dosud nepoznala. Kmeny v kladu CH13 / NeuroS1, kterému ICTV dosud nepřidělil druh, jsou označeny přerušovanou čarou. Hodnoty bootstrapu jsou uvedeny v uzlech. CH, Švýcarsko; JP, Japonsko; USA, Spojené státy americké; UY, Uruguay; GB, Spojené království Velké Británie; DE, Německo; SE, Švédsko; FR, Francie; CN, Čína. BoAstV, bovinní astrovirus; OvAstV, ovinní astrovirus; PoAstV, prasečí astrovirus; HuAstV, lidský astrovirus; MiAstV, norkový astrovirus.
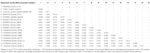
Tabulka 1. Odhady evoluční divergence na aminokyselinové úrovni kompletní ORF2 oblasti mezi sekvencemi skotu a ovcí Mamastrovirus-13 a kmeny v CH13/NeuroS1 clade nebyla zatím přiřazena k druhu Mezinárodní Výbor pro Taxonomii Virů (ICTV).
Studie založené na neuropatologické vyšetření a astrovirus nukleové kyseliny a bílkoviny zjištění k závěru, že existuje pravděpodobná příčinná souvislost mezi astrovirus infekce a neurologické onemocnění a lézí u skotu (Selimovic-Hamza et al., 2017a; Reuter a kol., 2018). Podle našich nejlepších znalostí nebyla encefalitida spojená s astrovirem experimentálně reprodukována. To by vyžadovalo izolaci neuroinvazivních astrovirů od klinických případů, což se v našem případě nepokusilo.
zdroj Chloubyv-Neuro-Uy v tomto případě nebylo možné určit. Je však třeba vzít v úvahu dobytek a volně žijící zvířata, protože skot byl chován za rozsáhlých venkovních podmínek. Postižené zvíře bylo zakoupeno a přesunuto na farmu v únoru 2018 spolu s dalšími 9 voly. Majitel bohužel odmítl další odběry a testování dalších zvířat v objektu a podrobnější epidemiologické šetření. U žádného z ostatních zvířat ve skupině nedošlo k neurologickému onemocnění od srpna 2018, Naposledy byl kontaktován Veterinární lékař. Sezónnost od začátku zimy do konce jara bylo navrženo pro případy astrovirus-spojené encefalitidy ve Švýcarsku (Selimovic-Hamza et al., 2016). Zajímavé je, že zde popsaný případ nastal v červnu, což odpovídá přechodnému období podzim-zima na jižní polokouli.
zatímco neurotropní astroviry byly identifikovány v Severní Americe (Li et al ., 2013; Spinato a kol., 2017), Evropa (Bouzalas et al., 2014) a Asie (Hirashima et al., 2018), jejich přítomnost nebyla nikdy zaznamenána na Jižní polokouli, takže tato komunikace rozšiřuje geografické rozložení astrovirus-spojené encefalitidy. Posoudit, zda virový kmen zjištěn v Uruguayi mají původ v Evropě, Severní Americe, nebo Asii, odhadli jsme, že evoluční rychlost a provedena analýza phylogeographic pomocí neuroinvasive BoAstV k dispozici sekvence v GenBank. Evoluční rychlost odhadovaná pomocí kompletní kódovací oblasti byla 4.27 × 10-4 (95% nejvyšší hustota pravděpodobnosti-HPD -, 2,19-6,46 × 10-4) substituce nukleotidů / místo / rok, což se očekává u RNA viru (Jenkins et al ., 2002), ale nižší, než se odhaduje u enterických lidských astrovirů (Babkin et al., 2012, 2014). Na ORF1ab regionu ukázaly podobné evoluční rychlost (4.20 × 10-4, 95% HPD 1.66–6.46 × 10-4 substituce/web/rok) jako kompletní kódování regionu, zatímco ORF1a (2.92 × 10-4, 95% HPD 1.19 × 10-6-6.46 × 10-4 substituce/web/rok) a ORF2 (2.86 × 10-4, 95% HPD 4.13 × 10-6-5.79 × 10-4 substituce/web/rok) vykazovaly mírně rychlejší evoluční rychlost, a částečné polymerázové genomické oblasti (ORF1b) vykazovaly mírně pomalejší evoluční rychlost (5.39 × 10-4, 95% HPD 6.41 × 10-7-1.10 × 10-3 substituce/web/rok).
Jak stanoví analýza phylogeographic s kompletní kódování regionu, a je znázorněno na MCTT (Obrázek 3), jsou tam dva sub-linie (CH13 a NeuroS1) na základě referenční kmeny, které mají společného předka. Poslední společný předek těchto sub-linie (lineage CH13/NeuroS1) vznikla v Evropě přibližně v roce 1885 (95% HPD, 1794-1940). Na začátku roku 1900, dva sub-linie rozešly, CH13 sub-linie zůstala v oběhu v Evropě, zatímco NeuroS1 sub-linie rozšířila do Ameriky a Asie. Nejpravděpodobnější scénář je, že NeuroS1 sub-rod byl představen v Uruguay z Evropy kolem roku 1921 (95% HPD, 1849-1967), pravděpodobně prostřednictvím hospodářských zvířat, obchodu, pak se rozšířil do Severní Ameriky a později do Japonska (viz Obrázek 3). Vzhledem k omezení v počtu sekvencí je k dispozici v GenBank, které by mohly mít neobjektivní analýzy, výsledky získané pomocí kompletní kódování regionu byly srovnávány s výsledky získanými s jinými genomových regionů (ORF1ab, ORF2, ORF1a, a ORF1b), k dispozici pro větší počet kmenů (tj. Kanadské kmeny). Ve všech analýzách je nejpravděpodobnějším scénářem, že k zavlečení viru do Uruguaye došlo z Evropy (doplňující údaje S1A-D). Kromě toho, předpokládané datum pro tento úvod, získané s ORF1ab a částečné polymerázové genomické oblasti (ORF1b) (Doplňující Údaje S1A,D), byl podobný jako získané s kompletní kódování regionu, vzhledem k tomu, že předpokládané datum zavedení získané s ORF2 a ORF1a bylo dříve, ale s širší 95% HPD intervalu (Doplňující Údaje, S1B,C). Úvod sub-linie NeuroS1 přímo do Kanady, z Evropy, s následným rozšířila do Spojených Států a Japonska, je také pravděpodobné, y zobrazeny na Doplňkovém Obrázku S1D.

obrázek 3. Maximální věrohodnost stromu clade získaná analýzou celovečerní kódovací oblasti. Barva pobočkami představuje největší pravděpodobností zemi, kde předkové oběhu, zadní pravděpodobnost hodnoty jsou uvedeny ve větvích a čísla v každém uzlu představují letech původu pro každé clade s 95% HPD intervalu. Podlinky jsou označeny štítky.
Další vyšetření jsou potřebné k posouzení geografické rozložení, patogenní mechanismy (zejména mechanismy přenosu a vstupu), molekulární epidemiologie, a potenciální mezidruhový přenos neuroinvasive astroviruses.
dostupnost dat
datové sady generované pro tuto studii lze nalézt v GenBank, MK386569.
autorské příspěvky
FG, RDC a MC přispěly koncepcí studie. FG a RDC provedly patologické vyšetření a odběr vzorků. PP provedl in situ hybridizaci. FU provedl imunohistochemii. LM, RC a MC provedly testování molekulární virologie. MC provedl sekvenční a fylogeografické analýzy a související obrázky. FG a PP získaly histologické obrazy. MF provedl bakteriální kultury. FG a MC napsali první návrh rukopisu. RDC, PP, FU, LM, MF, a RC napsal části rukopisu. Všichni autoři přispěli k revizi rukopisu, přečetli a schválili předloženou verzi.
finanční Prostředky
Tato práce byla financována Granty PL-015 N-15156 od INIA a 158 z „Programa de Iniciación a la Investigación 2017 „z“ Comisión Odvětvové de Investigación Científica “ (CSIC). MC a RDC potvrzují podporu „národní agentury pro výzkum a inovace“ (ANII) a INIA prostřednictvím Ph.D. stipendií. FG uznává podporu od ANII prostřednictvím mobility grant MOV_CA_2018_1_150021.
Prohlášení o střetu zájmů
autoři prohlašují, že výzkum byl proveden bez jakýchkoli obchodních nebo finančních vztahů, které by mohly být vykládány jako potenciální střet zájmů.
recenzent TS deklaroval minulé spoluautorství s jedním z autorů PP redakci manipulace.
Poděkování
autoři děkuji Yisell Perdomo a Cecilia Monesiglio od INIA, a Karen Sverlow a Juliann Beingesser z CAHFS pro technickou pomoc.
Doplňkový Materiál
Doplňkový Materiál pro tento článek lze nalézt online na: https://www.frontiersin.org/articles/10.3389/fmicb.2019.01240/full#supplementary-material
OBRÁZEK S1 | Maximální clade důvěryhodnost stromy (MCCTs) získané analýzou full-délka ORF1ab (A), full-délka ORF2 (B), full-délka ORF1a (C), a částečné ORF1b (D). Barva pobočkami představuje největší pravděpodobností zemi, kde předkové oběhu, zadní pravděpodobnost hodnoty jsou uvedeny ve větvích, a čísla v každém uzlu představují letech původu pro každé clade s 95% HPD intervalu. Podlinky jsou označeny štítky.
Poznámky pod čarou
- ^http://iqtree.cibiv.univie.ac.at
Al-Zoreky, N., a Sandine, W. E. (1990). Vysoce selektivní médium pro izolaci Listeria monocytogenes z potravy. Appl. Environ. Mikrobiol. 56, 3154–3157.
PubMed Abstraktní | Google Scholar
Ashbaugh, S. E., Thompsonová, K. E., Belknap, E. B., Schultheiss, P. C., Chowdhury, a. S., Collins, J. K. (1997). Specifická detekce vylučování a latence bovinního herpesviru 1 a 5 pomocí vnořené polymerázové řetězové reakce. J.Vet. Diagne. Investovat. 9, 387–394. doi: 10.1177/104063879700900408
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Babkin, I. V., Tikunov, v. A. Y., Sedelnikova, D. A., Zhirakovskaia, E. V., a Tikunova, N. V. (2014). Rekombinační analýza založená na kompletních genomech HAstV-2 a HastV-4. Infikovat. Genete. Evol. 22, 94–102. doi: 10.1016 / j. meegid.2014.01.010
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Babkin, I. V., Tikunov, v. A. Y., Zhirakovskaia, E. V., Netesov, S. V., a Tikunova, N. V. (2012). Vysoká evoluční rychlost lidského astroviru. Infikovat. Genete. Evol. 12, 435–442. doi: 10.1016 / j. meegid.2012.01.019
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Blomström, A. L., Widén, F., Kladivo, A. S., Belák, S., Berg, M. (2010). Detekce nového astroviru v mozkové tkáni norků trpících syndromem třepání norků pomocí virové metagenomiky. J. Clin. Mikrobiol. 48, 4392–4396. doi: 10.1128 / JCM.01040-10
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Boros,Á, Albert, M., Pankovics, P., Bíró, H., Pesavento, P. A., Phan, T. G., a kol. (2017). Ohniska neuroinvazivního astroviru spojená s encefalomyelitidou, slabost, a ochrnutí u odstavených prasat, Maďarsko. Emergu. Infikovat. Dis. 23, 1982–1993. doi: 10.3201/eid2312.170804
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Boujon, C. L. Koch, M. C., Wüthrich, D., Werder, S., Jakupovic, D., Bruggmann, R., et al. (2017). Indikace křížového přenosu astroviru spojeného s encefalitidou u ovcí a skotu. Emergu. Infikovat. Dis. 23, 1604–1608. doi: 10.3201 / eid2309.170168
PubMed Abstract / CrossRef Full Text / Google Scholar
Bouzalas, I. G., Zurthrich, D., Selimovic-Hamza, s., Zurscheller, C., Bruggmann, R., and Seuberlich, T. (2016). Plně genomově založené molekulární charakterizace astroviróz sdružených s encefalitidou skotu. Infect. Genete. Evol. 44, 162-168, 10.1016/J. meegid.2016.06.052
PubMed Abstract / CrossRef Full Text / Google Scholar
Bouzalas, I. G., Zurbriggen, a., Vandevelde, m., et al. (2014). Neurotropní astrovirus u skotu s nepodporující encefalitidou v Evropě. J. Clin. Mikrobiol. 52, 3318–3324. doi: 10.1128 / JCM.01195-14
PubMed Abstrakt / CrossRef Plný Text / Google Scholar
Cantile, C., and Youssef, s. (2016). „Nervový systém“, v Jubb, Kennedy, a Palmerova patologie domácích zvířat, 6. Edn, ed. M. G. Maxie (Saint Louis, MO: Elsevier), 205-406.
Google Scholar
Deiss, R., Selimovic-Hamza, S., Seuberlich, T., a Meylan, M (2017). Neurologické klinické příznaky u skotu s encefalitidou spojenou s astrovirem. J.Vet. Stážista. Med. 31, 1209–1214. doi: 10.1111/jvim.14728
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Donato, C., a Vijaykrishna, D. (2017). Široký rozsah hostitele a genetická rozmanitost savčích a ptačích astrovirů. Viry 9: E102. doi: 10.3390/v9050102
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Drummond, a. J. Suchard, M. a., Xie, D., a Rambaut, A. (2012). Bayesovská fylogenetika s krásou a šelmou 1.7. Molo. Biol. Evol. 29, 1969–1973. doi: 10.1093/molbev/mss075
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Giannitti, F., Anderson, M., Miller, M., Rowe, J., Sverlow, K., Vasquez, M., et al. (2016). Chlamydia pecorum: fetální a placentární léze u sporadických potratů koz. J.Vet. Diagne. Investovat. 28, 184–189. doi: 10.1177/1040638715625729
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Hirashima, Y., Okada, D., Shibata, S., Yoshida, S., Fujisono, S., Omatsu, T., et al. (2018). Analýza celého genomu nového neurotropního astroviru skotu zjištěného u japonského Černého vola s nehnisavou encefalomyelitidou v Japonsku. Oblouk. Virol. 163, 2805–2810. doi: 10.1007/s00705-018-3898-3
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Mezinárodní Výbor pro Taxonomii Virů (2018). Seznam hlavních druhů (MSL32), verze 1. Dostupné na adrese: https://talk.ictvonline.org/files/master-species-lists/m/msl/7185 (přístupné v březnu 2019).
Jenkins, G.M., Rambaut, a., Pybus, O. G., and Holmes, E. C. (2002). Míry molekulární evoluce RNA virů: kvantitativní fylogenetická analýza. J. Mol. Evol. 54, 156–165. doi: 10.1007/s00239-001-0064-3
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Kumar, S., Stecher, G., a Tamura, K. (2016). MEGA7: molecular evolutionary genetics analysis verze 7.0 pro větší datové sady. Molo. Biol. Evol. 33, 1870–1874. doi: 10.1093/molbev/msw054
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Li, L., Diab, S., McGraw, S., Barr, B., Traslavina, R., Higgins, R., et al. (2013). Divergentní astrovirus spojený s neurologickým onemocněním u skotu. Emergu. Infikovat. Dis. 19, 1385–1392. doi: 10.3201/eid1909.130682
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Lole, K. S., Bollinger, R. C., Paranjape, R. S., Gadkari, D., Kulkarni, S. S., Nováková, N. G., et al. (1999). Celovečerní virus lidské imunodeficience typu 1 genomy ze sérokonvertorů infikovaných subtypem C v Indii, s důkazem rekombinace intersubtypu. J. Virol. 73, 152–160.
PubMed Abstrakt / Google Scholar
Naccache, S. N., Peggs, K. S., Mattes, F. M., Phadke, R., Garson, J. a., Grant, P., et al. (2015). Diagnóza neuroinvazivní astrovirové infekce u imunokompromitovaného dospělého s encefalitidou nezaujatým sekvenováním nové generace. Cline. Infikovat. Dis. 60, 919–923. doi: 10.1093/cid/ciu912
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Palmieri, C., Franco, M., Uzal, F., Anderson, M., Barr, B., Woods, L., et al. (2011). Patologie a imunohistochemické nálezy infekce virem západního Nilu u psittaciformes. Veterinář. Patol. 48, 975–984. doi: 10.1177/0300985810391112
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Pfaff, F., Schlottau, K., Scholes, S., Courtenay, A., Hoffmann, B., Höper, D., et al. (2017). Nový astrovirus spojený s encefalitidou a ganglioneuritidou u domácích ovcí. Transbound. Emergu. Dis. 64, 677–682. doi: 10.1111 / tbed.12623
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Quan, P. L., Wagner, T. a., Briese, T., Torgerson, T. R., Narozdíl, M., Tashmukhamedova, A., et al. (2010). Astrovirus encefalitidy v chlapce s X-vázanou agamaglobulinémií. Emergu. Infikovat. Dis. 16, 918–925. doi: 10.3201/eid1606.091536
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Rambaut, A., Lam, T. T., Max Carvalho, L., a Pybus, O. G. (2016). Zkoumání časové struktury heterochronních sekvencí pomocí TempEst (formely Path-o-Gen). Virus Evol. 2: vew007. doi: 10.1093/ve/vew007
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Reuter, G., Pankovics, P., a Boros, A. (2018). Nepodporující (aseptická) meningoencefalomyelitida spojená s neurovirulentními astrovirovými infekcemi u lidí a zvířat. Cline. Mikrobiol. Rev. 31, e00040-18. doi: 10.1128 / CMR.00040-18
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Schlottau, K., Schulze, C., Düsseldorf, S., Hanke, D., Höper, D., Beer, M., et al. (2016). Detekce nového bovinního astroviru u krávy s encefalitidou. Transbound. Emergu. Dis. 63, 253–259. doi: 10.1111 / tbed.12493
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Selimovic-Hamza, S., Boujon, C. L., Hilbe, M., Oevermann, A., a Seuberlich, T. (2017a). Frekvence a patologický fenotyp skotu astrovirus CH13/NeuroS1 infekce u neurologicky-onemocnění skotu: k posouzení kauzality. Viry 9: E12. doi: 10.3390/v9010012
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Selimovic-Hamza, S., Sanchez, S., Philibert, H., Clark, E. G., a Seuberlich, T. (2017b). Infekce skotu astrovirem u skotu s neurologickým onemocněním v západní Kanadě. Si. Veterinář. J. 58, 601-603.
Google Scholar
Selimovic-Hamza, S., Bouzalas, I. G., v rámci vandevelde, M., Oevermann, A., a Seuberlich, T. (2016). Detekce astroviru v historických případech Evropské sporadické bovinní encefalitidy, Švýcarsko 1958-1976. Před. Veterinář. Věda. 3:91. doi: 10.3389 / fvets.2016.00091
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Seuberlich, T., Wüthrich, D., Selimovic-Hamza, S., Drögemüller, C., Oevermann, a., Bruggmann, R., et al. (2016). Identifikace druhého astroviru spojeného s encefalitidou u skotu. Emergu. Mikrobi Infikují. 5: 5. doi: 10.1038 / emi.2016.5
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Spinatu, M. T. Vince, A., Cai, H., a Ojkic, D. (2017). Identifikace bovinního astroviru v případech hnisavé encefalitidy skotu ve východní Kanadě. Si. Veterinář. J. 58, 607-609.
Google Scholar
Stein, L. T., Rech, R. R., Harrison, L., Brown, C. C. (2010). Imunohistochemická studie viru vztekliny v centrálním nervovém systému domácích a volně žijících druhů. Veterinář. Patol. 47, 630–633. doi: 10.1177/0300985810370013
PubMed Abstraktní | CrossRef Plný Text | Google Scholar
Trifinopoulos, J., Nguyen, L. T., von Haeseler, A., a Minh, B. Q. (2016). W-IQ-TREE: rychlý online fylogenetický nástroj pro analýzu maximální pravděpodobnosti. Nukleové Kyseliny Res.44, W232–W235. doi: 10.1093/nar / gkw256
PubMed Abstrakt / CrossRef Plný Text / Google Scholar
Tse, h., Chan, W. m., Tsoi, h. W., Fan, R. Y., Lau, C. C., Lau, s. K., et al. (2011). Znovuobjevení a genomická charakterizace bovinních astrovirů. J. Gen. Virol. 92 (Pt 8), 1888-1998. doi: 10.1099 / vir.0.030817-0
PubMed Abstrakt / CrossRef Plný Text / Google Scholar