Zellbiologie 06: Das Zytoskelett Teil II: Tubulin
Dies sind Notizen aus Vorlesung 6 des Cell Biology Course der Harvard University.
Letzte Woche haben wir Mikrofilamente behandelt, die aus Aktin bestehen. Diese Woche: Mikrotubuli aus Tubulin. Warum, fragen Sie, braucht das Zytoskelett zwei getrennte Systeme? Bedenken Sie, dass Mikrofilamente nur Ketten einzelner Untereinheiten sind, während Mikrotubuli buchstäblich Röhren sind (wie wir in Kürze sehen werden) – Hohlzylinder mit Wänden aus Ketten aus Dimeren. Diese unterschiedlichen Strukturen bedeuten unterschiedliche Fähigkeiten: Mikrofilamente können sich leichter spontan bilden und sich verzweigen (mit Hilfe von Arp2 / 3 und anderen zuletzt diskutierten Komplexen), um unterschiedliche Netzwerkformen zu bilden und verschiedene Teile der Zelle zu verbinden. Mikrotubuli verlassen sich stärker auf die Keimbildung, um sich zu bilden, und sind im Grunde ein zelluläres Netzwerk, das das Zentrosom mit der Peripherie der Zelle verbindet. In der Praxis sind Mikrofilamente im Zellkortex reichlich vorhanden und stark an kontraktilen Bewegungen und Zellmotilität beteiligt, während Mikrotubuli am stärksten an der Organisation von Organellen beteiligt sind und als Spuren für den anterograden und retrograden Transport dienen.
Mikrotubuli
( Bild dank Wikimedia Commons (Jeffrey81)
Neben Wikipedia sind auch frühere Ausgaben der auf NCBI verfügbaren Lehrbücher zur Zellbiologie von Lodish und Cooper hervorragende Referenzen.
Die röhrenförmige Struktur von Mikrotubuli ist stabiler als Mikrofilamente und ermöglicht viel schweres Ziehen und Schieben. Obwohl robust, sind Mikrotubuli temperaturabhängig – sie depolymerisieren, wenn sie auf 4 ° C abgekühlt werden, und werden wieder repolymerisieren, wenn sie auf 37 ° C erhitzt werden, vorausgesetzt, dass GTP verfügbar ist.
Der Grundbaustein der Mikrotubuli ist ein α-Tubuilin/β-Tubulin-Dimer (kodiert durch TUBA_-Gene bzw. Die einzelnen Tubulinproteine wiegen jeweils ~ 55kDa, also bei ~ 110Da / Aminosäure in der Größenordnung von 500 Aminosäuren. Sowohl α- als auch β-Tubulin binden GTP, aber α hält es so ziemlich für immer fest, während es in β ausgetauscht oder zu GDP hydrolysiert und dann wieder gegen neues GTP ausgetauscht werden kann.
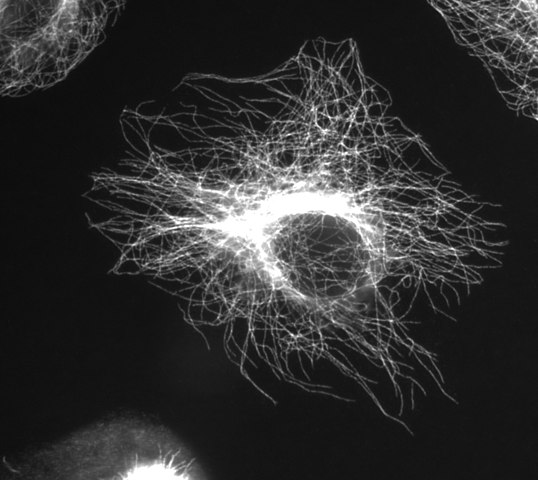
Stränge aufeinanderfolgender α / β-Dimere bilden Protofilamente, und 13 Protofilamente, die nebeneinander in einem Zylinder angeordnet sind, bilden ein Mikrotubuli. Der Raum zwischen den Protofilamenten wird als Naht bezeichnet. Der gesamte Mikrotubulus hat einen Durchmesser von ~ 25 nm. Sie können seine Struktur (aus Dimeren und Protofilamenten) im Innenleben des Zellsegments über Mikrotubuli sehen:
Innerhalb jedes Dimers ist die β-Untereinheit das für die Polymerisation bevorzugte (+) Ende und das α das für die Depolymerisation bevorzugte (-) Ende. Mikrotubuli bilden sich im Wesentlichen in den gleichen drei Schritten wie Mikrofilamente – Keimbildung, Dehnung und Steady State. Aber im Gegensatz zu Mikrofilamenten keimen sie nicht leicht von selbst, so dass die Keimbildung Mikrotubuli-Organisationszentren (MTOCs) erfordert. Nicht teilende Zellen haben jeweils nur ein MTOC, das Zentrosom genannt wird. (Dies ist auch im Video oben dargestellt). Das Zentrosom befindet sich in der Nähe des Zellkerns und unterliegt einer radialen Konfiguration von Mikrotubuli mit (-) Enden, die nach innen zeigen, und (+) Enden, die zur Zellperipherie zeigen.
Das Zentrosom besteht aus 2 Zylindern, den Zentriolen. Jede Zentriole besteht aus jeweils 9 Sätzen von 3 lateral fusionierten Mikrotubuli, die von einem amorphen ‚perizentriolären Material‘ umgeben sind, das reich an Dingen ist, die die Keimbildung fördern – insbesondere γ-Tubulin-Ringkomplexe (Gamma-Tubulin wird von den TUBG_-Genen kodiert). γ-Tubulin wird als ‚Split Washer‘ betrachtet:

Das Modell ist, dass die gespaltenen Enden γ-Tubulin erlauben, an α-Tubulin – d.h. das (-) Ende eines zu bildenden Mikrotubulus – zu binden, das den Keimbildungssamen für die Bildung eines Mikrotubulus bereitstellt.
Mikrotubuli können in vitro gebildet werden. Die Dynamik hängt hauptsächlich von kritischen Konzentrationen ab. Das (-) Ende ist weniger zur Polymerisation geneigt als das (+) Ende, daher hat es eine höhere kritische Konzentration. Wenn die tatsächliche Konzentration zwischen den kritischen Konzentrationen der beiden Enden liegt, tritt Tretmühle auf.
Wenn ein Mikrotubuli plötzlich zu dissoziieren beginnt, spricht man von einer ‚Katastrophe‘. Katastrophen haben eine interessante energetische Dynamik. Denken Sie daran, dass Beta-Tubulin, das das (+) Ende ist, an dem die Dehnung stattfindet, entweder GTP- oder GDP-gebunden sein kann. Es ist ein stabilerer Teil seines Mikrotubulus, wenn es GTP-gebunden ist; BIP-gebundenes Tubulin neigt zur Dissoziation. Mikrotubuli bilden sich hauptsächlich durch Zugabe von GTP-gebundenem Beta-Tubulin am (+) -Ende, aber nach Zugabe hydrolysieren die Beta-Tubulinmoleküle später ihr GTP und lassen sie GDP-gebunden. Es gibt also eine Art ‚Spitze‘ des Mikrotubulus, die GTP-gebunden ist, während Beta-Tubulin, das tiefer im Mikrotubulus liegt und vor längerer Zeit hinzugefügt wurde, GDP-gebunden ist. Wenn die Grenze der GTP-Hydrolyse die Spitze einholt oder wenn etwas passiert, um den Mikrotubulus zu durchtrennen, dann wird das weniger stabile, GDP-gebundene Beta-Tubulin freigelegt, und die Protofilamente beginnen sich wie „Widderhörner“ abzuschälen. In diesem Fall zerlegt sich der Mikrotubulus, bis er irgendwo weiter unten im Stroh auf eine „Insel“ aus GTP-gebundenem Beta-Tubulin trifft. Die Katastrophe kann durch die Zugabe neuer GTP-gebundener Tubulindimere, die die Enden abdecken und die Protofilamente stabilisieren, ‚gerettet‘ werden, so dass sich die Mikrotubuli neu bilden können. Sie können in diesem Video sehen, dass die Depolymerisation von Mikrotubuli viel schneller sein kann als die Polymerisation:
Hier sind einige Verbindungen, die für die Untersuchung von Mikrotubuli nützlich sind. Colchicin, ein Medikament gegen Gicht, bindet freie Alpha-Beta-Dimere, reduziert deren Zufuhr zur Mikrotubulibildung und fördert so die Depolymerisation. Nocodazol stört auch die Bildung neuer Mikrotubuli, und da die Bildung neuer Mikrotubuli für die Mitose wichtig ist, ist Nocodazol ein antineoplastisches Krebsmedikament. Umgekehrt wirkt Paclitaxel (Taxol), ein weiteres Krebsmedikament, durch Stabilisierung von Mikrotubuli, da der Abbau vorhandener Mikrotubuli auch für die Mitose wichtig ist.
Mikrotubuli-assoziierte Proteine (MAPs) haben verschiedene Rollen. Bemerkenswert sind MAP4 (in nicht-neuronalen Zellen), MAP2 (in Neuronen) und Tau (in Neuronen; kodiert durch MAPT-Gen). Allen gemeinsam ist, dass sie Mikrotubuli stabilisieren und die Kinetik zugunsten des Mikrotubuliwachstums und gegen die Katastrophe verschieben. Jedes enthält eine 18-Aminosäuregruppe mit positiv geladenen Aminosäuren, die an den negativen Teil von Mikrotubuli binden. Sie können mehrere Mikrotubuli bündeln , wobei MAP2 die Mikrotubuli aufgrund seines längeren Arms (im Vergleich zu Tau) in einem größeren Abstand hält.
Die Karten werden durch Phosphorylierung reguliert: die MARK-Proteinkinasen binden Phosphatgruppen kovalent an S-, T- oder Y-Aminosäuren in den MAP-Proteinen, wodurch ihre Fähigkeit, Mikrotubuli zu binden, verringert wird. (Cyclinabhängige Kinasen regulieren auch die MAPs während der Mitose). Es wird angenommen, dass diese Karten die physikalische Struktur von Zellen bestimmen. In Neuronen findet sich MAP2 in Dendriten und Tau hauptsächlich im Axon. Mutationen im MAPT-Gen, das für Tau kodiert, verursachen frontotemporale Demenz (FTD). Hyperphosphoryliertes Tau findet sich (obwohl wir nicht wissen warum) im Gehirn von Alzheimer-Patienten. Mausmodelle dieser beiden Krankheiten zeigen eine axonale Degeneration , die damit übereinstimmt, dass Tau seine Aufgabe, Mikrotubuli im Axon zu stabilisieren, nicht erfüllen kann.
Eine andere Klasse von Proteinen, die wegen ihrer Bindung an das (+) Ende von Mikrotubuli +Tips genannt wird, kann vor einer Katastrophe schützen. Sie scheinen ein paar Wege nach unten in die Mikrotubuli zu verlängern. Dieses Video zeigt, wie EB-1 nach außen gedrückt wird, weil die Enden der Mikrotubuli wachsen:
Andere endbindende Proteine fördern eher die Stabilität als die Stabilität. Kinesin-13 wirkt, um das Ende der Protofilamente zu krümmen und die Schwelle für eine Katastrophe zu senken. Stathmin (STMN1-Gen; auch bekannt als Op18, wobei Op für Onkoprotein steht) bindet an Tubulindimere innerhalb eines Protofilaments und fördert sowohl die sofortige Katastrophe als auch möglicherweise auch die GTP-Hydrolyse, die durch Entfernen von „Inseln“ von GTP-gebundenem Beta-Tubulin eher eine langfristige Investition zugunsten der Katastrophe darstellen würde. Stathmin wird durch Phosphorylierung reguliert. Katanin (nach dem japanischen Schwert Katana) durchtrennt buchstäblich Mikrotubuli.
Mikrotubuli-Motorproteine sind den Mikrofilament-Motorproteinen weitgehend ähnlich. Sie kommen in zwei Familien: Kinesine, von denen sich die meisten anterograde bewegen, dh zum (+) Ende; und Dyneine, von denen sich die meisten retrograd bewegen, dh zum (-) Ende. Sie ‚gehen‘ so:
Kinesin und Dynein sind an der Bewegung von Organellen, Endozytose und Exozytose und Chromosomensegregation während der Meiose beteiligt & Mitose.
Kinesin-1, das am besten untersuchte der Kinesine, ist insofern ein ‚konventionelles‘ Kinesin, als es als Tetramer aus zwei schweren Ketteneinheiten (KIF1-Gene z. KIF1A), die die ‚Kopf‘-Domänen umfassen, die ATP hydrolysieren und die Mikrotubuli binden, und zwei leichte Ketteneinheiten (KLC-Gene, z. B. KLC1), die einen ‚Schwanz‘ umfassen, der die Ladung bindet, in diesem Fall Vesikel. Eine Linkerdomäne (zu welchem Protein gehört dieses?) ermöglicht die Dimerisierung von zwei schweren Ketten.
Hier ist ein Video davon in Aktion. Beachten Sie, dass das Video es ein Dimer anstelle eines Tetramers nennt; Ich denke, das liegt daran, dass sie nur die Köpfe betrachten und nicht die Schwänze diskutieren.
Einige der anderen Kinesins unterscheiden sich ein wenig:
- Kinesin-2, das auch Vesikel & Organellen transportiert, ist ein Heterotrimer mit zwei verschiedenen (wenn auch verwandten) schweren Ketten und einem anderen Polypeptid, mit dem es seine Ladung reguliert.
- Kinesin-5 hat an beiden Enden Köpfe anstelle von Kopf und Schwanz, so dass es in entgegengesetzte Richtungen auf zwei verschiedenen Mikrotubuli läuft und sie zusammenzieht. Das nennt man bipolare Bewegung.
- Kinesin-14 ist das einzige bekannte Kinesin, das sich zum (-) Ende hin bewegt und an der Mitose beteiligt ist.
Die Prinzipien der Gehbewegung sind jedoch für alle Kinesine ziemlich gleich und werden im letzten Video oben dargestellt. Wenn sie nicht an einen Mikrotubulus gebunden sind, sind die ‚Köpfe‘ beide ADP-gebunden. Ein Kopf wird zufällig auf einen Mikrotubulus treffen und daran binden, sein ADP freisetzen, so dass ein ATP es ersetzen kann. Die ATP-Bindung induziert Konformationsänderung, die auf dem Linker zieht, Schwingen die anderen, ‚hinken‘ Kopf nach vorne in die führende Position, wo es an den Mikrotubuli bindet. Der ursprüngliche Kopf hydrolysiert dann ATP, das Phosphat freisetzt (im Video als Pi abgekürzt) und die Energie für den einen energetisch aktiven Schritt in diesem Prozess liefert, der sich vom Mikrotubulus löst.
Myosin (das Motorprotein, das auf Mikrofilamenten läuft) und Kinesin haben sehr ähnliche Strukturen, aber keine Ähnlichkeit der Aminosäuresequenz. Daher wird angenommen, dass sie keine Paraloge sind, sondern ein Beispiel für konvergente Evolution.
Die Leute reden viel über Kinesine, zum Teil, weil wir Dyneine nicht so gut verstehen. Dyneine sind riesige Proteine, die aus 2 großen, 2 mittleren und 2 kleinen Untereinheiten bestehen. Ihre schiere Größe hat es schwierig gemacht, sie zu isolieren und zu charakterisieren, und ihre Funktionsweise ist nicht gut verstanden. Wir wissen, dass der Dynaktinkomplex (ein Multiproteinkomplex; die Dynaktine selbst sind die DTCN_-Gene) als ‚Adapter‘ beteiligt ist, der Dynein mit Fracht verbindet.
Dieses Video fasst das gesamte Zytoskelett zusammen und enthält einen Großteil des Materials von letzter Woche und dieser Vorlesung:
PrP
Wie in den Secretory Pathway Notes erwähnt, ist PrP GPI-verankert an der Membran und wird sehr regelmäßig Endozytose unterzogen, wodurch endozytäre Vesikel mit PrP in ihnen entstehen. Encalada 2011 stellt fest, dass Kinesin-1C und DHC1 (Dynein Heavy Chain 1) für den Transport dieser PrP-Vesikel anterograde bzw. retrograde verantwortlich sind. Wenn Encalada ausgeschlagen, niedergeschlagen oder Kinesin-1C gehemmt wurde, waren anterograde und retrograde Bewegungen reduziert, und in ähnlicher Weise, wenn DHC1 gestört war. Es scheint also, dass diese beiden Komplexe, obwohl sie sich in entgegengesetzte Richtungen bewegen, sich gegenseitig aktivieren. Interessanterweise verhinderte die Störung dieser Motorproteine nicht, dass PrP-Vesikel mit Motoren in Verbindung gebracht wurden – sie bewegten sich einfach nicht so schnell oder so oft. Das Papier hat eine Reihe anderer zellbiologischer Schlussfolgerungen darüber, wie Vesikel Motorproteine aktivieren und wie die Transportrichtung bestimmt wird.