biologia Pääaineilla I
oppimistulokset
- ymmärtää esi-mRNA: n ja mRNA: n eron
transkription jälkeen eukaryoottiset pre-mrnat on käytävä läpi useita käsittelyvaiheita ennen kuin ne voidaan kääntää. Eukaryoottiset (ja prokaryoottiset) trnat ja rrnat käsitellään myös ennen kuin ne voivat toimia komponentteina proteiinisynteesiä valmistavissa koneissa.
mRNA-käsittely
eukaryoottista esi-mrnaa käsitellään laajasti ennen kuin se on valmis käännettäväksi. Eukaryoottisen mRNA: n kypsymiseen liittyvät lisävaiheet luovat molekyylin, jolla on huomattavasti pidempi puoliintumisaika kuin prokaryoottisella mRNA: lla. Eukaryoottiset mrnat kestävät useita tunteja, kun taas tyypillinen E. coli mRNA kestää enintään viisi sekuntia.
esi-mRNA: t päällystetään ensin RNA: ta stabiloivilla proteiineilla; nämä suojaavat esi-mRNA: ta hajoamiselta, kun sitä käsitellään ja viedään ulos tumasta. Kolme tärkeintä vaihetta ennen mRNA-käsittelyä ovat stabilointi-ja signalointikertoimien lisääminen molekyylin 5′ ja 3′ päissä sekä niiden välissä olevien sekvenssien poistaminen, jotka eivät määritä sopivia aminohappoja. Harvoissa tapauksissa mRNA: n transkriptio voidaan ”muokata” sen jälkeen, kun se on litteroitu.
5 ’yläraja
esi-mRNA: ta edelleen syntetisoitaessa lisätään 7-metyyliguanosiinikorkki kasvavan transkription 5′ päähän fosfaattiliitoksella. Tämä osa (funktionaalinen ryhmä) suojaa orastavaa mRNA: ta hajoamiselta. Lisäksi proteiinisynteesiin osallistuvat tekijät tunnistavat korkin ribosomien käännöksen aloittamiseksi.
3 ’ Poly-a-pyrstö
kun venymä on valmis, esi-mRNA halkaistaan endonukleaasilla AAAAAA-konsensusjonon ja gu-rikkajonon välillä, jolloin AAAAAA-sekvenssi jää esi-mRNA: lle. Poly-a-polymeraasi-niminen entsyymi lisää tämän jälkeen noin 200 A: n jäämiä sisältävän narun, jota kutsutaan poly-a-pyrstöksi. Tämä muutos suojaa edelleen esi-mRNA: ta hajoamiselta ja viestii transkription tarvitsemien solutekijöiden viennistä sytoplasmaan.
mRNA: ta edeltävät geenit
eukaryoottiset geenit koostuvat eksoneista, jotka vastaavat proteiinia koodaavia sekvenssejä (ex-on tarkoittaa, että ne ilmaistaan), ja interventiosekvensseistä, joita kutsutaan introneiksi (introniksi, joka tarkoittaa niiden väliroolia), jotka voivat osallistua geenien säätelyyn, mutta jotka poistetaan esi-mRNA: sta käsittelyn aikana. MRNA: n intronijaksot eivät koodaa funktionaalisia proteiineja.
intronien löytyminen tuli yllätyksenä 1970-luvulla tutkijoille, jotka olettivat, että ennen mrnoja määritettäisiin proteiinisekvenssejä ilman jatkokäsittelyä, kuten he olivat havainneet prokaryooteilla. Korkeampien eukaryoottien geeneissä on hyvin usein yksi tai useampia introneja. Nämä alueet voivat vastata säätelysekvenssejä, mutta sen biologinen merkitys, Että geenissä on useita introneja tai hyvin pitkiä introneja, on epäselvä. On mahdollista, että intronit hidastavat geeniekspressiota, koska esi-mrnojen transkriptiointi runsailla introneilla kestää kauemmin. Vaihtoehtoisesti intronit voivat olla epäfunktionaalisia sekvenssijäänteitä, jotka ovat jääneet jäljelle muinaisten geenien fuusioitumisesta koko evoluution ajan. Tätä tukee se, että erilliset eksonit koodaavat usein erillisiä proteiinialayksiköitä tai domeeneja. Suurimmaksi osaksi intronien sekvenssit voivat mutatoitua vaikuttamatta lopulta proteiinituotteeseen.
kaikki mRNA: ta edeltävät intronit on poistettava kokonaan ja tarkasti ennen proteiinisynteesiä. Jos prosessi erehtyy yksikin nukleotidi, uudelleen yhdistettyjen eksonien lukukehys muuttuisi ja syntyvä proteiini olisi toimintahäiriöinen. Prosessia, jossa intronit poistetaan ja eksonit yhdistetään uudelleen, kutsutaan liitokseksi (Kuva 1). Intronit poistuvat ja hajoavat esi-mRNA: n ollessa vielä tumassa. Liitos tapahtuu sekvenssispesifisellä mekanismilla, joka varmistaa, että intronit poistuvat ja eksonit yhdistyvät uudelleen yhden nukleotidin tarkkuudella. Pre-mRNAs: n liittäminen tapahtuu proteiini-ja RNA-molekyylien komplekseilla, joita kutsutaan spliseosomeiksi.
käytännön kysymys
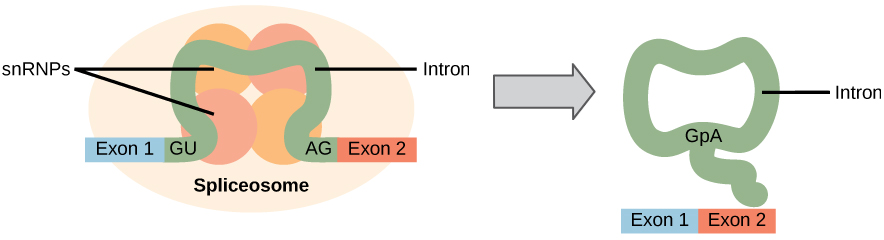
Kuva 1. MRNA: ta edeltävässä liitoksessa intronit poistetaan tarkasti primäärisestä RNA-transkriptiosta. Splikaatioprosessia katalysoivat reliseosomeiksi kutsutut proteiinikompleksit, jotka koostuvat snrnas-nimisistä proteiineista ja RNA-molekyyleistä. Spliceosomit tunnistavat sekvenssejä intronin 5′ ja 3 ’ päässä.
liitosvirheet liittyvät syöpiin ja muihin ihmisten sairauksiin. Millaiset mutaatiot voivat johtaa liitosvirheisiin?
huomaa, että yli 70 yksittäistä intronia voi olla läsnä, ja jokainen on läpäistävä prosessi—lisäksi 5′ rajaus ja lisäämällä poly-a hännän—vain tuottaa yhden, käännettävissä mRNA molekyyli.
RNA: n muokkaus Trypanosomeissa
kuva 2. Trypanosoma brucei on unitaudin aiheuttaja ihmisillä. Tämän taudinaiheuttajan mRNAs-arvoja on muutettava nukleotideja lisäämällä ennen proteiinisynteesiä. (luotto: Torsten Ochsenreiterin teoksen muokkaus)
trypanosomit ovat alkueläinten ryhmä, johon kuuluu ihmisellä unitautia aiheuttava taudinaiheuttaja Trypanosoma brucei (kuva 2). Trypanosomeissa ja käytännössä kaikissa muissakin eukaryooteissa on mitokondrioiksi kutsuttuja organelleja, jotka toimittavat solulle kemiallista energiaa. Mitokondriot ovat omaa DNA: taan ilmentäviä organelleja, joiden uskotaan olevan jäänteitä eukaryootin ja nielaistun prokaryootin symbioottisesta suhteesta. Trypanosomien mitokondrio-DNA: ssa on mielenkiintoinen poikkeus keskeiseen Opinkappaleeseen: niiden esi-mrnoilla ei ole oikeaa tietoa funktionaalisen proteiinin määrittelemiseksi. Yleensä tämä johtuu siitä, että mRNA: sta puuttuu useita U-nukleotideja. Solu suorittaa ylimääräisen RNA-prosessointivaiheen, jota kutsutaan RNA-muokkaukseksi tämän korjaamiseksi.
mitokondrion genomin muut geenit koodaavat 40 – 80-nukleotidisopastetta RNAs. Yksi tai useampi näistä molekyyleistä vuorovaikuttaa komplementaarisella emäspariutumisella joidenkin mRNA: ta edeltävän transkription nukleotidien kanssa. Opas-RNA: ssa on kuitenkin enemmän nukleotideja kuin esi-mRNA: ssa on U-nukleotideja, joihin sitoutua. Näillä alueilla opas-RNA loopsahtaa ulos. Guide RNAs: n 3 ’ -päissä on pitkä poly-U-pyrstö, ja nämä U-emäkset lisätään Pre-mRNA-transkription alueille, joissa opas Rnat on silmukoitu. Tämä prosessi on täysin RNA-molekyylien välittämä. Toisin sanoen RNA: n muokkauksessa katalyytteinä toimivat pikemminkin ohjeelliset RNA: t kuin proteiinit.
RNA-editointi ei ole vain trypanosomien ilmiö. Joidenkin kasvien mitokondrioissa muokkautuvat lähes kaikki esi-mrnat. RNA: n muokkausta on havaittu myös nisäkkäillä, kuten rotilla, kaneilla ja jopa ihmisillä. Mikä voisi olla evolutiivinen syy tähän mRNA: ta edeltävään lisäaskeleeseen? Yksi mahdollisuus on, että mitokondrioilla, jotka ovat muinaisten prokaryoottien jäänteitä, on yhtä vanha RNA-pohjainen menetelmä geenien ilmentymisen säätelyyn. Tämän hypoteesin tueksi mrnoja edeltäviksi tehdyt muokkaukset vaihtelevat soluolosuhteiden mukaan. Vaikka RNA: n muokkaus on spekulatiivista, se saattaa jäädä alkutaipaleelta, jolloin RNA-molekyylit proteiinien sijaan olivat vastuussa reaktioiden katalysoinnista.
Kokeile.
Osallistu!
paranna tätä sivua lisää