biologia Pääaineille II
oppimistulokset
- ymmärtää aiemmat ja uudet virusten luokittelujärjestelmät
koska useimmat virukset ovat todennäköisesti kehittyneet eri esivanhemmista, tutkijoiden käyttämät systemaattiset menetelmät prokaryoottisten ja eukaryoottisten solujen luokittelemiseksi eivät ole kovin hyödyllisiä. Jos virukset edustavat eri eliöiden ”jäänteitä”, ei edes genomi-tai proteiinianalyysistä ole hyötyä. Miksi?, Koska viruksilla ei ole yhteistä genomisekvenssiä, jonka ne kaikki jakavat. Esimerkiksi 16S rRNA-sekvenssi, joka on niin hyödyllinen prokaryoottifylogenioiden rakentamisessa, ei hyödytä olentoa, jolla ei ole ribosomeja! Biologit ovat aiemmin käyttäneet useita luokittelujärjestelmiä. Virukset ryhmiteltiin aluksi jaetun morfologian mukaan. Myöhemmin virusten ryhmät luokiteltiin niiden sisältämän nukleiinihapon, DNA: n tai RNA: n tyypin mukaan ja sen mukaan, oliko niiden nukleiinihappo yksi – vai kaksijuosteinen. Nämä aikaisemmat luokittelumenetelmät ryhmittelivät virukset kuitenkin eri tavalla, koska ne perustuivat viruksen eri merkistöihin. Nykyisin yleisimmin käytettyä luokittelumenetelmää kutsutaan Baltimoren luokittelujärjestelmäksi, ja se perustuu siihen, miten lähetti-RNA (mRNA) syntyy kussakin tietyssä virustyypissä.
aiemmat luokittelujärjestelmät
virukset sisältävät vain muutamia elementtejä, joiden perusteella ne voidaan luokitella: viruksen perimä, kapsidin tyyppi ja vaipallisten virusten kuorirakenne. Kaikkia näitä alkuaineita on käytetty aiemmin virusluokituksessa (Taulukko 1 ja kuva 1). Viruksen genomit voivat vaihdella geneettisen materiaalin tyypin (DNA tai RNA) ja sen organisaation (yksi – tai kaksijuosteinen, lineaarinen tai pyöreä, ja segmentoitu tai segmentoimaton) mukaan. Joissakin viruksissa replikaatioon tarvittavat lisäproteiinit liittyvät suoraan genomiin tai sisältyvät viruksen kapsidiin.
| Taulukko 1. Viruksen luokittelu genomin rakenteen ja ytimen mukaan | |
|---|---|
| Ydinluokitukset | esimerkkejä |
| RNA | rabiesvirus, retrovirukset |
| DNA | herpesvirukset, isorokkovirus |
| yksijuosteiset | rabiesvirus, retrovirukset |
| kaksijuosteiset | herpesvirukset, isorokkovirus |
| lineaariset | rabiesvirus, retrovirukset, herpesvirukset, isorokkovirus |
| Kiertokirje | papilloomavirukset, monet bakteriofagit |
| segmentoimaton: genomi koostuu yhdestä geeniainesosasta | Parainfluenssaviruksia |
| segmentoitu: genomi jakautuu useisiin segmentteihin | influenssavirukset |
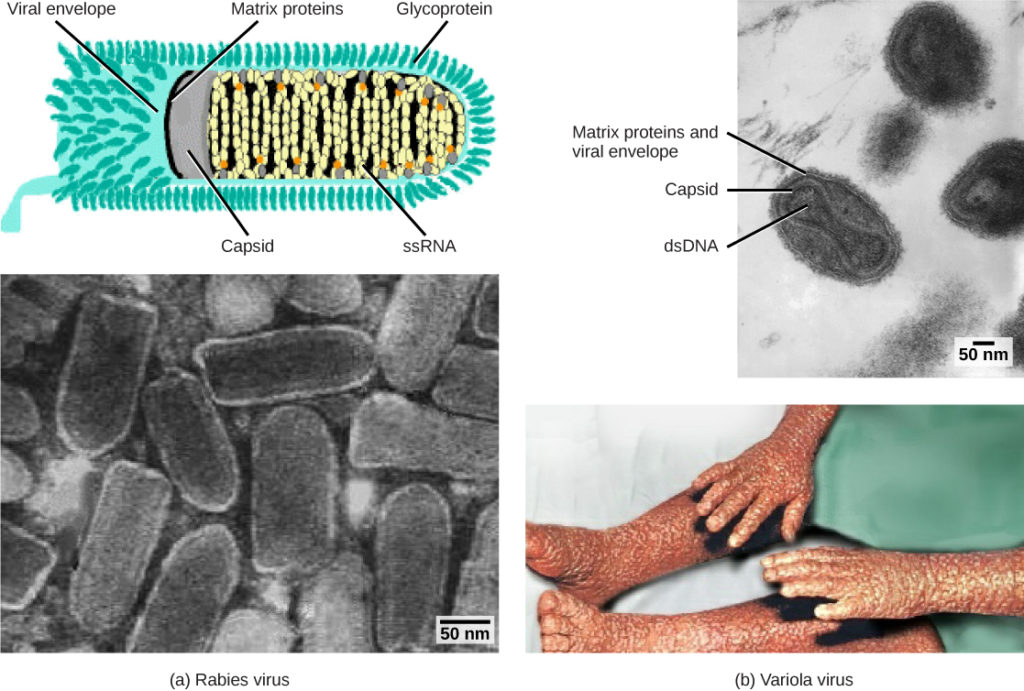
Kuva 1. Virukset luokitellaan niiden perimän ja kapsidin rakenteen perusteella. a) Rabiesviruksella on yksijuosteinen RNA (ssRNA)-ydin ja vaipallinen kierteinen kapsidi, kun taas B) isorokkoa aiheuttavalla variola-viruksella on kaksijuosteinen DNA (dsDNA) – ydin ja monimutkainen kapsidi. (luotto ”rabies diagram”: modification of work by CDC; ”rabies micrograph”: modification of work by Dr. Fred Murphy, CDC; luotto ”small pox micrograph”: modification of work by Dr. Fred Murphy, Sylvia Whitfield, CDC; luotto ”isorokko photo”: modification of work by CDC; mittakaavapalkin tiedot Matt Russellilta)
virukset voidaan luokitella myös niiden kapsidien rakenteen perusteella (Taulukko 2 ja kuva 2). Kapsidit luokitellaan alastomiin ikosaedrisiin, vaipallisiin ikosaedrisiin, vaipallisiin kierteisiin, alastomiin kierteisiin ja kompleksisiin. Viruksen ydinrakenteiden luokittelussa käytetään geneettisen materiaalin tyyppiä (DNA tai RNA) ja sen rakennetta (yksi – tai kaksijuosteinen, lineaarinen tai pyöreä sekä segmentoitu tai segmentoimaton) (Taulukko 2).
| Taulukko 2. Viruksen luokittelu kapsidin rakenteen mukaan | |
|---|---|
| Kapsidiluokitus | esimerkkejä |
| alaston ikosaedrinen | hepatiitti A-virus, poliovirukset |
| vaipallinen Ikosaedrinen | Epstein-Barr-virus, herpes simplex-virus, vihurirokkovirus, keltakuumevirus, HIV-1 |
| vaipallinen kierteinen | influenssavirukset, sikotautivirukset, tuhkarokkovirukset, raivotautivirukset |
| paljas kierteinen | tupakan mosaiikkivirus |
| monimutkainen monien proteiinien kanssa; joillakin on ikosaedristen ja kierteisten kapsidirakenteiden yhdistelmiä | herpesvirukset, isorokkovirus, hepatiitti B-virus, T4-bakteriofagi |

kuva 2. Eri virusten transmissioelektronimikrografit näyttävät niiden rakenteet. A) polioviruksen kapsidi on paljas ikosaedrinen; B) Epstein-Barr-viruksen kapsidi on vaipallinen ikosaedrinen; c) sikotautiviruksen kapsidi on vaipallinen kierteinen; d) tupakan mosaiikkiviruksen kapsidi on paljas kierteinen; ja e) herpesviruksen kapsidi on monimutkainen. (credit a: modification of work by Dr. Fred Murphy, Sylvia Whitfield; credit b: modification of work by Liza Gross; credit c: modification of work by Dr. F. A. Murphy, CDC; credit d: modification of work by USDA ARS; credit e: modification of work by Linda Stannard, Department of Medical Microbiology, University of Cape Town, South Africa, NASA; scale-bar data from Matt Russell)
Baltimoren luokitus
yleisimmin käytetyn virusluokitusjärjestelmän kehitti Nobel-palkittu biologi David Baltimore 1970-luvun alussa. edellä mainittujen morfologian ja genetiikan erojen lisäksi Baltimoren luokittelujärjestelmä ryhmittelee virukset sen mukaan, miten mRNA syntyy viruksen replikaatiosyklin aikana.
I-ryhmän virukset sisältävät perimänään kaksijuosteista DNA: ta (dsDNA). Niiden mRNA: ta tuotetaan transkriptiolla paljolti samalla tavalla kuin solujen DNA: ta.
ryhmän II viruksilla on perimänään yksijuosteinen DNA (ssDNA). Ne muuntavat yksijuosteiset genominsa dsDNA-välituotteeksi ennen kuin transkriptio mRNA: han voi tapahtua.
III ryhmän virukset käyttävät genominaan dsRNA: ta. Säikeet erkanevat toisistaan, ja yhtä niistä käytetään mallina mRNA: n generoinnissa viruksen koodaaman RNA-riippuvaisen RNA-polymeraasin avulla.
ryhmän IV viruksilla on genominaan ssRNA, jonka polariteetti on positiivinen. Positiivinen polaarisuus tarkoittaa, että genominen RNA voi toimia suoraan mRNA: na. DsRNA: n välituotteita, joita kutsutaan replikatiivisiksi välituotteiksi, valmistetaan kopioimalla genomista RNA: ta. Näistä välituotteista muodostuu useita, täyspitkiä RNA-säikeitä, joilla on negatiivinen polaarisuus (jotka täydentävät positiivisjuosteista genomi-RNA: ta), jotka voivat sitten toimia malleina RNA: n tuottamiseksi positiivisella polaarisuudella, mukaan lukien sekä täyspitkät genomiset RNA: t että lyhyemmät virus-mrnat.
V-ryhmän virukset sisältävät ssRNA-genomeja, joiden polaarisuus on negatiivinen, eli niiden sekvenssi täydentää mRNA: ta. Kuten ryhmän IV viruksilla, dsRNA-välituotteita käytetään genomin kopioimiseen ja mRNA: n tuottamiseen. Tällöin negatiivissäikeinen genomi voidaan muuntaa suoraan mRNA: ksi. Lisäksi täyspitkistä positiivisista RNA-säikeistä tehdään malleja negatiivisjuosteisen genomin tuottamiseksi.
ryhmän VI viruksilla on diploidi (kaksi kopiota) ssRNA-genomi, joka on muunnettava käänteiskopioijaentsyymin avulla dsDNA: ksi; dsDNA kulkeutuu isäntäsolun tumaan ja siirtyy isäntäsolun perimään. Tämän jälkeen mRNA: ta voidaan tuottaa transkriptiolla viruksen DNA: sta, joka oli integroitu isäntägenomiin.
ryhmän VII viruksilla on osittainen dsDNA-genomi ja ne tekevät ssRNA-välituotteita, jotka toimivat mRNA: na, mutta muuntuvat myös takaisin dsDNA-genomeiksi käänteiskopioijaentsyymin avulla, mikä on tarpeen genomin replikoinnissa. Baltimoren luokituksen kunkin ryhmän ominaisuudet on koottu taulukossa 3 kunkin ryhmän esimerkkeihin.
| Taulukko 3. Baltimoren luokitus | |||
|---|---|---|---|
| ryhmä | ominaisuudet | mRNA: n tuotantotapa | esimerkki |
| I | kaksijuosteinen DNA | mRNA on litteroitu suoraan DNA-templaatista | Herpes simplex (herpesvirus) |
| II | yksijuosteinen DNA | DNA muuttuu kaksijuosteiseksi ennen kuin RNA transkriboidaan | Koiran parvovirus (parvovirus) |
| III | kaksijuosteinen RNA | mRNA on transkriboitu RNA-genomi | lapsuuden gastroenteriitti (rotavirus) |
| IV | yksijuosteinen RNA (+) | genomi toimii mRNA: na | tavallinen flunssa (pircornavirus) |
| V | yksijuosteinen RNA (−) | mRNA on transkriboitu RNA-genomista | Rabies (rabdovirus) |
| VI | yksijuosteiset RNA-virukset, joiden käänteiskopioijaentsyymi on | Käänteiskopioijaentsyymi muodostaa DNA: ta RNA-genomista; DNA on sitten sisällytetty isäntägenomiin; mRNA on transkriboitu yhdistetystä DNA: sta | ihmisen immuunikatovirus (HIV) |
| VII | kaksijuosteiset DNA-virukset, joiden käänteiskopioijaentsyymi on | viruksen perimä on kaksijuosteinen DNA, mutta viruksen DNA replikoituu RNA: n välivaiheen kautta; RNA voi toimia suoraan mRNA: na tai mallina mRNA: n tekemiseksi | hepatiitti B-virus (hepadnavirus) |