Frontiers in Microbiology
Introduction
Astroviridae-heimo sisältää vaipattomia, positiivi-aistisia, yksijuosteisia RNA-viruksia kahdessa suvussa, Mamastroviruksessa ja avastroviruksessa, jotka tartuttavat nisäkkäitä ja lintuja. Tällä hetkellä International Committee on Taxonomy of Viruses (International Committee on Taxonomy of Viruses , 2018) tunnustaa 19 lajia, eli Mamastrovirus-1: stä -19: ään, Mamastrovirus-suvun sisällä; on kuitenkin olemassa lukuisia kantoja, jotka odottavat luokittelua, joista osaa pidetään alustavasti uusina lajeina (Donato ja Vijaykrishna, 2017).
vuodesta 2010 lähtien useat astrovirukset on yhä useammin tunnustettu neuroinvasiivisiksi eri nisäkäslajeilla, myös ihmisillä (Quan et al., 2010; Naccache et al., 2015), minkki (Blomström ym., 2010), karja (Li et al., 2013), sheep (Pfaff et al., 2017), ja sikoja (Boros et al., 2017). Naudan astrovirukseen liittyvän enkefaliitin alustavan toteamisen jälkeen yhdysvaltalaiskarjassa (Li et al., 2013), retrospektiivinen tutkimus satunnaisen naudan enkefaliitin tapauksissa määrittelemätön etiologia Sveitsistä paljasti, että tämä neuroinvasive astrovirus oli mennyt huomaamatta vuosikymmeniä (Selimovic-Hamza et al., 2016). Vaikka näiden astrovirusten epidemiologiaa ja tarttumisreittejä ei tunneta, lajien välistä tarttumista on ehdotettu perustuen nauta-ja lammasperäisten neuroinvasitiivisten astrovirusten suureen identiteettiin (>98%) nukleotidi-ja aminohappotasoilla (Boujon et al., 2017).
naudan astrovirukset (BoAstVs), nimeltään BoAstV-NeuroS1 (Li et al., 2013)ja BoAstV-CH13 (Bouzalas et al., 2014), löydettiin alun perin non-suppuratiivista enkefaliittia sairastavien nautojen aivoista Yhdysvalloissa ja Sveitsissä. Eri nimikkeistöstä huolimatta molemmat virukset edustavat samaa genotyyppilajia (Bouzalas et al., 2016; Selimovic-Hamza et al., 2017a), joka odottaa vielä ICTV: n virallista luokitusta. Vuonna 2015 Sveitsissä havaittiin aiemmin tuntematon BoAstV-kanta, jonka nimi on BoAstV-CH15, lehmien aivotulehdusta sairastavien lehmien aivoista. Koko genomin fylogeneettinen vertailu paljasti boastv-CH15: n läheisemmän suhteen lampaan astrovirukseen (OvAstV) kuin BoAstV-CH13: een (Seuberlich et al., 2016). Koinfektio boastv-CH13: n ja BoAstV-CH15: n kanssa dokumentoitiin myös yhdessä tapauksessa (Seuberlich et al., 2016). Samana vuonna Saksassa, Schlottau et al. (2016) kertoi uudesta astroviruksesta, nimittäin boastv-BH89/14, lehmässä, jolla on enkefaliitti, joka oli läheisintä sukua OvAstV: lle ja BoAstV-CH15: lle. Myöhemmin BoAstV-CH13 / NeuroS1 todettiin vuonna 2017 naudan enkefaliittitapauksissa itä-ja Länsi-Kanadassa (Spinato et al., 2017; Selimovic-Hamza et al., 2017b). Vuonna 2018, uusi neuroinvasive BoAstV läheistä sukua Pohjois-Amerikan ja Euroopan Boastv-NeuroS1/BoAstV-CH13, tunnistettiin härkä ei-märkivä enkefalomyeliitti Japanissa, ja esiintyminen genotyyppisen rekombinaation Pohjois-Amerikan ja Euroopan kantoja ehdotettiin (Hirashima et al., 2018).
vaikka astrovirukseen liittyvää enkefaliittia on raportoitu Pohjois-Amerikassa, Euroopassa ja Aasiassa, niiden esiintymistä ei ole koskaan dokumentoitu eteläisellä pallonpuoliskolla. Tässä kuvataan astrovirukseen liittyvän enkefaliitin tapaus Uruguayn karjassa, joka laajentaa neuroinvasiivisten astrovirusten maantieteellistä levinneisyyttä ja geneettistä monimuotoisuutta ja tarjoaa fylogeografisia todisteita, jotka viittaavat siihen, että tämä virus tuotiin Amerikkoihin Euroopasta ja myöhemmin levisi Aasiaan.
materiaalit ja menetelmät
historia ja merkinanto
kesäkuussa 2018 22-kuukautinen Holstein-härkä 37 härän ryhmässä 300 hehtaarin maatilalla Coloniassa, Uruguayssa, kehittyi eteneväksi neurologiseksi lauluksi, johon kuului epätavallinen käyttäytyminen, päämäärätön kävely, kiertoliike, ataksia, toistuvat ja koordinoimattomat kielen liikkeet ja taantuminen. Lauma laidunsi jokavuotisella kauralaitumella ja sitä täydennettiin maissisäilörehulla. Eläinlääkärin oletettu kliininen diagnoosi aivojen listerioosista johti penisilliini-ja streptomysiinihoitoon, mutta eläin kuoli spontaanisti 3 päivää kestäneen kliinisen kuurin jälkeen.
patologiset tutkimukset, in situ hybridisaatio (Ish) ja immunohistokemia (IHC)
härän pää poistettiin ruhosta ja toimitettiin inia: n eläinlääketieteelliseen diagnostiseen laboratorioon (Animal Health Platform) diagnostista työtä varten. Puolet aivoista, lyhyt osa proksimaalista kohdunkaulan selkäydintä (C1), kolmoishermostoa ja kolmoishermon juurta, sylkirauhanen, nielun imusolmuke, nielu, ruokatorvi, kieli ja luurankolihas, upotettiin 10-prosenttisesti neutraaliin puskuroituun formaliiniin 48-72 tunnin ajan. kudoksia käsiteltiin rutiininomaisesti histologiaa varten, ne upotettiin parafiiniin, mikrotomi-osastoitiin 4-5 µm ja värjättiin hematoksyliinilla ja eosiinilla (H&E) ja Grammatahroilla.
kromogeeninen ISH tehtiin manuaalisesti 5 µm: n osalle formaliinikiinnitettyä, parafiinisesti upotettua (FFPE) aivo -, aivo-ja pikkuaivoja Superfrost Plus-dioilla (Thermo Fisher Scientific, Pittsburgh, PA, Yhdysvallat) käyttäen RNAscope 2.5 Red assay kit-testisarjaa (Cat #322360, Advanced Cell Diagnostics, Hayward, CA, Yhdysvallat) ja BoAstV probe Cat-koettinta. #406921. Luotain koostuu 20zz-pareista, jotka kohdistuvat alueelle 5232-6180 virusta (GenBank KF233994. 1). Jokainen 5 µm: n kudososa esikäsiteltiin kuumuudella ja proteaasilla ennen koettimen hybridisaatiota 2 tunnin ajan 40°C: ssa ja käsiteltiin valmistajan suositusten mukaisesti. Signaalin validoinnissa käytettyjä negatiivisia kontrolleja olivat muun muassa toisiinsa liittymätön (GC-pitoisuus sovitettu) koetin, joka suoritetaan sarjalohkoilla, ja tartuttamattomien eläinten kudokset. Dioja torjuttiin hematoksyliinilla ja asennettiin Ecomountilla (Biocare Medical, Concord, CA, Yhdysvallat).
lisäksi IHC: tä tehtiin aiemmin kuvatulla tavalla BRAINSTEMIN, aivojen ja pikkuaivojen FFPE-osissa Länsi-Niilin viruksen (WNV, Flavivirus) tunnistamiseksi (Palmieri et al., 2011), rabiesvirus (lyssavirus) (Stein et al., 2010)ja klamydia spp. (Giannitti et al., 2016) antigeenit.
Molekyylivirologia
nukleiinihapon uutto suoritettiin jäädytettyjen (-20°C) aivojen yhdistetystä näytteestä MagMAX Nucleic Acid Isolation Kit® – menetelmällä (Thermo Fisher Scientific). Astroviruksen toteamiseksi reverse transkriptio (RT) suoritettiin RevertAid-Käänteiskopioijaentsyymillä® (Thermo Fisher Scientific) ja satunnaisilla heksameerin alukkeilla (Qiagen). PCR tehtiin cDNA: sta käyttäen MangoMix®: tä (Bioliini) ja alukkeita, jotka vahvistavat astroviruksen polymeraasigeenin 432-nukleotidifragmenttia (Tse et al., 2011). PCR-tuote visualisoitiin 2% agaroosigeelissä, joka puhdistettiin PureLink® Quick Gel Extraction and PCR Purification Combo Kit (Invitrogen) – valmisteella ja sekvensoitiin Macrogen Inc. Soul, Etelä-Korea). Astroviruksen koko genomin amplifikaatio, Maxima H miinus Käänteiskopioijaentsyymi(Thermo Fisher Scientific) ja oligo (DT)18 cDNA: n obtentioon ja MangoMix® (Bioline) tai Ranger DNA polymeraasi (Bioline) hirashiman et al.kuvaamilla alukkeilla., 2018, käytettiin. PCR-tuotteet visualisoitiin 1-2-prosenttisessa agaroosigeelissä, joka puhdistettiin ja sekvensoitiin edellä mainitulla tavalla. Sekvenssi kokoonpano suoritettiin SeqMan (Lasergene 8, DNASTAR). Kaksikymmentäkuusi täydellistä genomisekvenssiä neuroinvasive astrovirus naudoista, lampaista, sioista, ihmisistä ja minkeistä, ja enteric naudan astrovirus saatavilla GenBank ladattiin ja kohdistettiin käyttäen Clustal W MEGA 7 ohjelmisto (Kumar et al., 2016). W-IQ-TREE1 (Trifinopoulos et al., 2016) käytettiin määrittämään parhaiten sopiva sekvenssievoluution malli (SYM+I+G4) ja rakentamaan maksimitodennäköinen fylogeneettinen puu, jossa on tässä tapauksessa havaitut BoAstV: n lähes täydelliset sekvenssit ja genbankista ladatut täydelliset sekvenssit käyttäen Bootstrapia tilastollisena menetelmänä kladien kestävyyden arvioimiseksi. Samankaltaisuus juoni suoritettiin SimPlot software (Lole et al., 1999). P-etäisyydet ORF2: n aminohappotasolla arvioitiin MEGA 7-ohjelmistolla (Kumar et al., 2016).
lisäksi BEAST v1.8.4-paketilla tehtiin Bayesilainen fylogeografinen analyysi (Drummond et al., 2012), käyttäen: täydellinen koodausalue BoAstV CH13 / NeuroS1 lineage, ORF1ab (ei-rakenteelliset geenit), ORF2 (rakenteellinen geeni), orf1a (proteaasi) ja osittainen ORF1b (polymeraasi genomialue, jolle Kanadan kantoja oli saatavilla), kaikki sekvenssit saatavilla GenBank (viimeinen liittyminen Huhtikuu 18, 2019), määrittää evolutiivinen nopeus, ikä/vuotta yhteisiä esivanhempia, ja todennäköisin reitti viruskiertoon maittain (Sveitsi, Uruguay, Yhdysvallat, Kanada, ja Japani). Rekombinaation puute aineistossa määritettiin rekombinaation Toteamisohjelmalla 4. Korvausmalli, joka parhaiten sopii kuhunkin kohdistukseen, määritettiin MEGA 7-ohjelmiston avulla Bayesilaisen informaatiokriteerin (BIC) avulla, ja kunkin aineiston ajallinen rakenne arvioitiin Tempestin (Rambaut et al., 2016). Lognormaali rento molekyylikello bayesilaisella Skyline-analyysillä valittiin Bayes Factor-menetelmällä käytettyjen molekyylikellojen ja koalitioisten puukierrosten erilaisten yhdistelmien joukosta. Tunnistusmaata käytettiin piirteenä. Markovin ketjun Monte Carlon pituus oli 100 miljoonaa sukupolvea, mikä varmisti Tracer v1.6: ssa arvioidun analyysin lähentymisen.0, ja posteriorista todennäköisyyttä käytettiin kladien arviointiin. Maximum clade uskottavuus tree (MCCT) saatiin TREEANNOTATOR ohjelmisto BEAST ja visualisoidaan Figtree v1.4.3.
lopuksi PCR käsitteli jäätyneistä aivoista uutetun DNA: n naudan herpesvirusten 1 ja 5 (BHV-1 ja -5) toteamiseksi edellä kuvatulla tavalla (Ashbaugh et al., 1997).
bakteriologia
tuoreita näytteitä aivoista ja aivoista käsiteltiin rutiininomaisesti aerobisia bakteeriviljelmiä varten blood-ja MacConkey-agar-bakteereissa sekä selektiivisiä Listeria monocytogenes-bakteeriviljelmiä varten (Al-Zoreky and Sandine, 1990).
tulokset ja keskustelu
tässä kuvatun tapauksen kliiniset oireet ja epidemiologiset löydökset, vaikkakin epäspesifiset, olivat samanlaiset kuin muissa naudan astrovirukseen liittyvän enkefaliitin tapauksissa, joita kuvataan yleensä sporadisina (Selimovic-Hamza ym., 2016), jossa on erilaisia neurologisia alijäämiä (Deiss et al., 2017), jonka kliinisten oireiden kesto vaihtelee tyypillisesti 1 päivästä 3 viikkoon (Schlottau et al., 2016; Deiss et al., 2017; Spinato et al., 2017; Hirashima et al., 2018).
aivojen, selkäytimen C1-segmentin ja muiden pään kudosten makroskooppisessa tutkimuksessa ei havaittu merkittäviä vakavia anatomisia leesioita. Histologisesti, oli keskivaikea tai vaikea, lymfosyyttinen, histiocytic ja plasmacytic meningoencephalomyelitis vaikuttavat telencephalon (mukaan lukien aivopuolisko ja hippokampus), brainstem, ja ainoa tutkittu segmentti selkäytimen. Leesiot jakautuivat pääasiassa harmaaseen aineeseen ja rajoittaviin valkoisen aineen alueisiin. Hoidetuilla alueilla esiintyi perivaskulaarisia kalvosimia ja lymfoplasmasyyttisiä ja histiocyyttisiä tulehduksia sekä neuronaalinen nekroosi/neuronofagia ja viereisen neuropiilin glioosi. Sairastuneiden nekroottisten neuronien satellitoosi (luvut 1a–D). Vauriot olivat paljon harvinaisempia ja vakavia pikkuaivojen parenkymiassa, vaikka siellä oli multifokaalinen keskivaikea pikkuaivojen leptomeningiitti. H&E-ja Gram-tahroilla ei havaittu intralesionaalisia bakteereja. Muissa tutkituissa kudoksissa ei havaittu merkittäviä histologisia muutoksia.

Kuva 1. Histologiset leesiot aivorungossa (A, B) ja aivokuoressa (C,D) ja boastv-RNA: n havaitseminen aivokuoressa (E,G). Kuvat (A-D) ovat aivojen osia, joissa on H&E; kuvat E ja G ovat aivokuoren osia, jotka osoittavat hybridisaation kromogeenisen ISH: n avulla BoAstV-spesifisen koettimen avulla, vastapainona hematoksyliinillä; kuva F on aivokuoren dapb-koettimen sarjallinen osa (negatiivinen kontrolli), vastapainona hematoksyliinillä. (A) perivenulaarinen tila on merkittävästi laajentunut tulehdussolujen (enimmäkseen lymfosyyttien ja histiocytes), jotka myös infiltroituvat viereisen neuropil. (B) hermosolun keskellä on hypereosinofiilinen perikaryon ja karyorrhexis (nekroosi) ja hermosolu ympäröi lisääntynyt määrä gliasolujen (satellitoosi) ja tulehdussolujen. (C) leptomeninge on infiltroitunut lymfosyyttien ja histiocytes. D) aivokuoren alue, jossa on useita hypereosinofiilisiä (nekroottisia) neuroneja ja suuri alus, jossa on perivaskulaarinen lymfosyyttinen käsiraudat. (E) aivokuoren sarjamaisessa osassa runsas sytoplasmainen BoAstV-RNA-merkintä kuvataan voimakkaalla, rakeisella punaisella kromogeenikertymällä Soman ja neuronaalisten jatkeiden sytoplasmassa (E, G), jota ei esiinny hybridisaatiossa negatiivisen kontrollianturin (F) avulla.
keskushermoston histologisessa tutkimuksessa epäiltiin neuroinvasiivista virusinfektiota. Aivotulehdusta sairastavat naudat huolestuttavat, koska monet märehtijöiden neuropatogeenit ovat zoonoottisia (Cantile and Youssef, 2016); siten, diagnoosi enkefaliitti olisi kannustettava laaja laboratoriokokeet seuloa taudinaiheuttajia, kun se on mahdollista. Tässä kuvatussa tapauksessa IHCs WNV: n, rabiesviruksen ja klamydia spp: n osalta. BHV-1: n ja -5: n PCR: t olivat kaikki negatiivisia, eikä aivokudoksesta viljelty patogeenisiä bakteereja. Koska Härkä oli < 2 vuotta vanha eikä aivorungossa havaittu spongiformisia muutoksia, eläintä ei testattu naudan spongiformisen enkefalopatian (BSE) varalta, joka on täysikasvuisten nautojen eksoottinen tauti, jota ei ole koskaan raportoitu Uruguayssa. Lisäksi BSE ei ole tulehduksellinen (Cantile and Youssef, 2016).
in situ-hybridisaatio suoritettiin BoAstV-NeuroS1: stä tuotetulla koettimella, ja koettimen hybridisaatio oli runsasta sen sisällä ja rajoittui aivopuoliskon neuronien sytoplasmaan ja hippokampukseen (luvut 1e–g). Näillä alueilla koetin hybridisaatio kolokalisoituu nekroottisten neuronien ja glioosin alueiden kanssa, eikä koetin hybridisaatio ole havaittavissa gliasoluissa eli perivaskulaarisen hihansuun tulehdussoluissa. Pikkuaivoissa Ish ei havainnut virusnukleiinihappoa, sillä parenkyymissä oli vain vähäisiä tulehdusmuutoksia, mutta keskivaikea leptomeningiitti eli aivoinfarkti, mukaan lukien kohdat, joilla oli vaikea tulehdus. Tämä tarkoittaa, että Ish: n viruslevityksen toteaminen oli topografisesti rajallisempaa kuin enkefaliitti tutkituissa osissa, mitä on toisinaan kuvattu naudan BoAstV-CH13/NeuroS1-related encephalitis-tapauksissa (Selimovic-Hamza et al., 2017a, b). Syynä tähän satunnaiseen virus-RNA: n havaitsemisen puutteeseen lesisoiduilla aivojen alueilla saattaa olla Ish: n havaitsemisraja tai viruksen poistuminen tulehtuneilta aivoalueilta kuolinhetkeen mennessä, kuten aiemmin on esitetty (Selimovic-Hamza et al., 2017b). Ish ei odotetusti havainnut luotaimen hybridisaatiota negatiivisena kontrollina käytettävässä aivokudoksessa.
astrovirus havaittiin aivoissa RT-PCR: llä. Lähes täydellinen genomisekvenssianalyysi paljasti Mamastroviruskannan Ch13 / NeuroS1-kladissa, jonka nimesimme BoAstV-Neuro-Uy: ksi, sekvenssi talletettiin Genbankiin liittymisnumerolla MK386569. Fylogeneettinen analyysi paljasti läheisyyden muiden neuroinvasiivisten astrovirusten kanssa Virginia/Human-Mink-lammas (va / HMO) – kladissa (kuva 2), joka sisältää tunnetuimpia neuroinvasiivisia astroviruksia (Hirashima et al., 2018; Reuter ym., 2018). BoAstV-Neuro-Uy: n lähes täydellinen sekvenssi on pituudeltaan 6427 bp ja sen sekvenssidentiteetti on 94% kagoshimasr28-462-kannan kanssa. BoAstV-Neuro-Uy: n ominaisuudet ovat samanlaiset kuin muillakin sukulinjan CH13/NeuroS1 kannoilla: 5 ’ UTR-alue on 51 NT, orf1a (proteaasi) on 861 aminohappoa (aa), ORF1b on 523 aa (RNA-riippuvainen RNA-polymeraasi) ja ORF2 on 758 aa (kapsidiproteiini). Valitettavasti 3 ’ UTR: ää ei voitu sekvensoida, mutta poly(a) – pyrstön oletetaan olevan läsnä, koska oligo(dT)18: aa käytettiin cDNA: n saamiseksi. Lisäksi esiintyy heptamerinen AAAAAAC-sekvenssi, ribosomaalinen kehyksensiirtosignaali. P-etäisyydet ORF2: n aminohappotasolla vahvistivat tämän kannan yhdistämisen Ch13/NeuroS1-kladiin. P-etäisyydet < 0.35 välillä BoAstV-Neuro-Uy ja muut tämän kladin jäsenet (Taulukko 1) tukisi näiden viruskantojen luokittelua saman lajin sisällä; Mamastrovirus-13 on äskettäin ehdottanut muut kirjoittajat (Donato ja Vijaykrishna, 2017; Hirashima et al., 2018), joskin ICTV: n varma lajimääräys on vireillä. BoAstV-Neuro1: stä tuotetulla Ish-luotaimella oli 92,7% sekvenssidentiteetti BoAstV-Neuro-Uy: n kanssa.
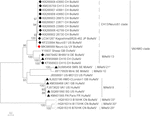
kuva 2. Täysipituisten nukleotidisekvenssien fylogeneettinen analyysi maksimitodennäköisyysmenetelmällä. BoAstV-Neuro-Uy on merkitty punaisella rhombuksella. Muut nautojen neuroinvasitiivisten astrovirusten sekvenssit on merkitty mustilla rhombeilla, muiden kuin nautojen neuroinvasitiivisten astrovirusten sekvenssit on merkitty mustilla kolmioilla ja enteeristen nautojen astrovirusten sekvenssit valkoisilla ympyröillä. Mamastroviruslajit on esitetty; tähdellä merkitään lajeja, joita ICTV ei ole vielä tunnistanut. Ch13/NeuroS1-kladin sisällä olevat kannat, joita ICTV ei ole vielä määrittänyt lajiksi, tunnistetaan katkaistulla viivalla. Bootstrap-arvot ilmoitetaan solmuissa. CH, Sveitsi; JP, Japani; US, Yhdysvallat; UY, Uruguay; GB, Ison-Britannian Yhdistynyt kuningaskunta; DE, Saksa; SE, Ruotsi; FR, ranska; CN, Kiina. BoAstV, naudan astrovirus; OvAstV, lampaan astrovirus; PoAstV, sian astrovirus; HuAstV, ihmisen astrovirus; MiAstV, minkin astrovirus.
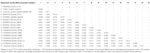
Taulukko 1. Arviot koko ORF2-alueen aminohappotason evoluutioeroista naudan ja lampaan Mamastrovirus-13-sekvenssien ja Ch13/NeuroS1-kladin kantojen välillä, joita kansainvälinen virusten Taksonomiakomitea (ICTV) ei ole vielä määrittänyt lajille.
neuropatologisiin tutkimuksiin ja astroviruksen nukleiinihapon ja proteiinin osoittamiseen perustuvissa tutkimuksissa on päätelty, että astrovirusinfektion ja nautojen neurologisen sairauden ja leesioiden välillä on todennäköinen syy-yhteys (Selimovic-Hamza et al., 2017a; Reuter et al., 2018). Tietääksemme astrovirukseen liittyvää aivotulehdusta ei ole vielä kokeellisesti uusittu. Tämä vaatisi neuroinvasiivisten astrovirusten eristämistä kliinisistä tapauksista, mitä ei meidän tapauksessamme yritetty.
BoAstV-Neuro-Uy: n lähdettä ei tässä tapauksessa voitu selvittää. Reserviläiskarja ja villieläimet kannattaa kuitenkin ottaa huomioon, sillä naudat kasvatettiin laajoissa ulkoilmaolosuhteissa. Sairastunut eläin oli ostettu ja siirretty tilalle helmikuussa 2018 yhdessä muiden 9 härän kanssa. Valitettavasti omistaja kieltäytyi ottamasta lisää näytteitä ja testaamasta muita eläimiä kiinteistössä sekä tarkemmasta epidemiologisesta tutkimuksesta. Yksikään ryhmän muista eläimistä ei ollut sairastunut neurologiseen sairauteen elokuuhun 2018 mennessä, jolloin eläinlääkäri oli viimeksi tavoitettu. Astrovirukseen liittyvän enkefaliitin tapauksille Sveitsissä on ehdotettu kausiluonteisuutta talven alusta kevään loppuun (Selimovic-Hamza et al., 2016). Mielenkiintoista on, että tässä kuvattu tapaus sattui kesäkuussa, mikä vastaa eteläisen pallonpuoliskon syys-talvi-siirtymäkautta.
kun taas neurotrooppisia astroviruksia on tunnistettu Pohjois-Amerikassa (Li et al., 2013; Spinato et al., 2017), Eurooppa (Bouzalas et al., 2014)ja Aasiassa (Hirashima et al., 2018), niiden esiintymistä ei ole koskaan raportoitu eteläisellä pallonpuoliskolla, joten tämä viestintä laajentaa astrovirukseen liittyvän enkefaliitin maantieteellistä levinneisyyttä. Arvioidaksemme, onko Uruguayssa havaittu viruskanta mahdollisesti peräisin Euroopasta, Pohjois-Amerikasta tai Aasiasta, arvioimme evoluutionopeuden ja teimme fylogeografisen analyysin neuroinvasive BoAstV-sekvensseillä, jotka ovat saatavilla Genbankissa. Koko koodausalueen avulla arvioitu evoluutionopeus oli 4.27 × 10-4 (95% suurin todennäköisyys tiheys-HPD-, 2.19–6.46 × 10-4) nukleotidisubstituutioita/paikka/vuosi, joka on odotettavissa RNA-viruksen (Jenkins et al., 2002), mutta pienempi kuin ihmisen enteristen astrovirusten arvioitu arvo (Babkin et al., 2012, 2014). ORF1ab-alueella oli samanlainen evoluutionopeus (4, 20 × 10-4, 95% HPD 1, 66–6, 46 × 10-4 substituutiota / paikka / vuosi) kuin koko koodausalueella, kun taas ORF1a (2, 92 × 10-4, 95% HPD 1, 19 × 10-6, 46 × 10-4 substituutiota/paikka/vuosi) ja ORF2 (2, 86 × 10-4, 95% HPD 4, 13 × 10-6-5.79 × 10-4 substituutiota/paikka/vuosi) oli hieman nopeampi evoluutionopeus, ja osittainen polymeraasin genomialue (ORF1b) osoitti hieman hitaampaa evoluutionopeutta (5, 39 × 10-4, 95% HPD 6, 41 × 10-7-1, 10 × 10-3 substituutiota/paikka/vuosi).
fylogeografisessa analyysissä, jossa on täydellinen koodausalue, ja joka on esitetty MCTT: ssä (kuva 3), on kaksi vertailukantoihin perustuvaa alalinjaa (CH13 ja NeuroS1), joilla on yhteinen esi-isä. Näiden alalinjojen viimeisin yhteinen esi-isä (sukujuuret CH13 / NeuroS1) syntyi Euroopassa noin vuonna 1885 (95% HPD, 1794-1940). 1900-luvun alussa nämä kaksi alalinjaa erosivat toisistaan, Ch13-alalinja jäi kiertämään Euroopassa, kun taas NeuroS1-alalinja levisi Amerikkaan ja Aasiaan. Todennäköisin skenaario on, että NeuroS1-alalinja otettiin käyttöön Uruguayssa Euroopasta noin vuonna 1921 (95% HPD, 1849-1967), oletettavasti karjakaupan kautta, sitten levisi Pohjois-Amerikkaan ja myöhemmin Japaniin (kuva 3). Koska genbankissa saatavilla olevien sekvenssien määrä rajoittui, mikä olisi voinut haitata analyysiä, tuloksia, jotka saatiin käyttämällä täydellistä koodausaluetta, verrattiin muihin genomialueisiin (ORF1ab, ORF2, ORF1a ja ORF1b), jotka olivat saatavilla suuremmalle määrälle kantoja (eli kanadalaisia kantoja). Kaikissa analyyseissä todennäköisin skenaario on, että viruksen kulkeutuminen Uruguayhin tapahtui Euroopasta (täydentävät luvut S1A–D). Lisäksi arvioitu käyttöönottopäivä, joka saatiin orf1ab: llä ja osittaisella polymeraasigenomialueella (Orf1b) (Lisäluvut S1A,D), oli samanlainen kuin koko koodausalueella, kun taas ORF2: lla ja ORF1a: lla saatu arvioitu käyttöönottopäivä oli aikaisempi,mutta laajempi 95 prosentin HPD-väli (Lisäluvut S1B, C). Käyttöönotto alalinjan NeuroS1 suoraan Kanadaan Euroopasta, myöhemmin levinnyt Yhdysvaltoihin ja Japaniin, on myös uskottava, s esitetty täydentävä Kuva S1D.

kuva 3. Suurin clade uskottavuus puu saadaan analysoimalla kokopitkä koodaus alue. Haarojen väri edustaa todennäköisintä maata, jossa esi-isät kiertävät, posterioriset todennäköisyysarvot esitetään haaroissa ja numerot kussakin solmussa kuvaavat kunkin kladin alkuperävuosia 95% HPD-välillä. Alalinjat on merkitty etiketeillä.
lisätutkimuksia tarvitaan neuroinvasiivisten astrovirusten maantieteellisen levinneisyyden, patogeenisten mekanismien (erityisesti tartunta-ja sisäänmenomekanismien), molekyyliepidemiologian ja mahdollisen lajien välisen leviämisen arvioimiseksi.
tietojen saatavuus
tätä tutkimusta varten luodut aineistot löytyvät genbankista, MK386569.
Tekijäosuudet
FG, RDC ja MC osallistuivat tutkimuksen suunnitteluun. FG ja RDC suorittivat patologisen tutkimuksen ja näytteenoton. PP suoritti in situ-hybridisaation. FU suoritti immunohistokemian. Lm, RC ja MC suorittivat molekyylivirologian testejä. MC suoritti sekvenssi-ja fylogeografiset analyysit ja niihin liittyvät luvut. FG ja PP saivat histologiset kuvat. MF suoritti bakteeriviljelmät. FG ja MC kirjoittivat käsikirjoituksen ensimmäisen luonnoksen. RDC, PP, FU, LM, MF ja RC kirjoittivat osia käsikirjoituksesta. Kaikki kirjoittajat osallistuivat käsikirjoituksen tarkistamiseen, lukivat ja hyväksyivät toimitetun version.
Rahoitus
tämä työ rahoitettiin INIAN myöntämillä apurahoilla PL-015 N-15156 ja ”Comisión Sectorial de Investigación Científican ”(CSIC)” Programa de iniciación a la Investigación 2017″. MC ja RDC tunnustavat tuen ”National Agency for Research and Innovation” (Anii) ja INIA, vastaavasti, kautta Ph. D. stipendejä. FG myöntää tukea anii: lta liikkuvuusapurahalla MOV_CA_2018_1_150021.
Eturistiriitalausunto
kirjoittajat toteavat, että tutkimus tehtiin ilman kaupallisia tai taloudellisia suhteita, joita voitaisiin pitää mahdollisena eturistiriitana.
arvostelija TS julisti aiemmin kirjoittajaksi yhden kirjoittajista PP: n käsittelevälle toimittajalle.
kiitokset
kirjoittajat kiittävät YISELL Perdomoa ja Cecilia Monesigliota INIASTA sekä Karen Sverlow ’ ta ja Juliann Beingesseriä cahfs: stä teknisestä avusta.
Täydennysaineisto
tämän artikkelin Täydennysaineisto löytyy verkosta osoitteesta: https://www.frontiersin.org/articles/10.3389/fmicb.2019.01240/full#supplementary-material
kuva S1 / Maximum clade-uskottavuuspuut (MCCTs), jotka on saatu analysoimalla kokopitkä ORF1ab (a), täyspitkä ORF2 (B), täyspitkä ORF1a (C) ja osittainen ORF1b (D). Haarojen väri edustaa todennäköisintä maata, jossa esi-isät kiertävät, posterioriset todennäköisyysarvot esitetään haaroissa ja numerot kussakin solmussa edustavat kunkin kladin alkuperävuosia 95% HPD-välillä. Alalinjat on merkitty etiketeillä.
alaviitteet
- ^http://iqtree.cibiv.univie.ac.at
Al-Zoreky, N., and Sandine, W. E. (1990). Erittäin selektiivinen kasvualusta Listeria monocytogenes-bakteerin eristämiseen elintarvikkeista. Appl. Environ. Mikrobiolia. 56, 3154–3157.
PubMed Abstract / Google Scholar
Ashbaugh, S. E., Thompson, K. E., Belknap, E. B., Schulteiss, P. C., Chowdhury, S., and Collins, J. K. (1997). Naudan herpesviruksen 1 ja 5 erityismääritys sisäkkäisellä polymeraasiketjureaktiolla. J. Vet. Diagn. Sijoittaa. 9, 387–394. doi: 10.1177/104063879700900408
PubMed Abstract | CrossRef Full Text | Google Scholar
Babkin, I. V., Tikunov, A. Y., Sedelnikova, D. A., Žirakovskaja, E. V., and Tikunova, N. V. (2014). Rekombinaatioanalyysi perustuu hastv-2: n ja hastv-4: n täydellisiin genomeihin. Tartuttaa. Genet. Evol. 22, 94–102. doi: 10.1016 / j.meegid.2014.01.010
PubMed Abstract | CrossRef Full Text | Google Scholar
Babkin, I. V., Tikunov, A. Y., Žirakovskaja, E. V., Netesov, S. V., and Tikunova, N. V. (2012). Ihmisen astroviruksen nopea evoluutio. Tartuttaa. Genet. Evol. 12, 435–442. doi: 10.1016 / j.meegid.2012.01.019
PubMed Abstract | CrossRef Full Text | Google Scholar
Blomström, A. L., Widén, F., Hammer, A. S., Belák, S., and Berg, M. (2010). Uuden astroviruksen osoittaminen ravistelevan minkin oireyhtymästä kärsivän minkin aivokudoksesta viruksen metagenomiikan avulla. J. Clin. Mikrobiolia. 48, 4392–4396. doi: 10.1128 / JCM.01040-10
PubMed Abstract | CrossRef Full Text | Google Scholar
Boros, Á, Albert, M., Pankovics, P., Bíró, H., Pesavento, P. A., Phan, T. G., et al. (2017). Neuroinvasiivisen astroviruksen taudinpurkaukset, jotka liittyvät enkefalomyeliittiin, heikkouteen ja halvaantumiseen vieroitettujen sikojen keskuudessa Unkarissa. Emerg. Tartuttaa. Tämä. 23, 1982–1993. doi: 10.3201 / eid2312.170804
PubMed Abstract | CrossRef Full Text | Google Scholar
Boujon, C. L., Koch, M. C., Wüthrich, D., Werder, S., Jakupovic, D., Bruggmann, R., et al. (2017). Tiedot enkefaliittiin liittyvän astroviruksen lajien välisestä tartunnasta lampaissa ja naudoissa. Emerg. Tartuttaa. Tämä. 23, 1604–1608. doi: 10.3201 / eid2309.170168
PubMed Abstract / CrossRef Full Text / Google Scholar
Bouzalas, I. G., Wüthrich, D., Selimovic-Hamza, S., Drögemüller, C., Bruggmann, R., and Seuberlich, T. (2016). Enkefaliittiin liittyvien nautaeläinten astrovirusten täydellinen genomipohjainen molekylaarinen karakterisointi. Tartuttaa. Genet. Evol. 44, 162-168. doi: 10.1016 / J. meegid.06/052/2016
PubMed Abstract / CrossRef Full Text / Google Scholar
Bouzalas, I. G., Wüthrich, D., Walland, J., Drögemüller, C., Zurbriggen, A., Vandevelde, M., et al. (2014). Neurotrooppinen astrovirus nautojen nonsuppurative enkefaliitti Euroopassa. J. Clin. Mikrobiolia. 52, 3318–3324. doi: 10.1128 / JCM.01195-14
PubMed Abstract | CrossRef Full Text | Google Scholar
Cantile, C., and Youssef, S. (2016). ”Nervous System” teoksessa Jubb, Kennedy, and Palmer ’ s Pathology of Domestic Animals, 6th Edn, toim. M. G. Maxie (Saint Louis, MO: Elsevier), 205-406.
Google Scholar
Deiss, R., Selimovic-Hamza, S., Seuberlich, T., and Meylan, M. (2017). Astrovirukseen liittyvää enkefaliittia sairastavien nautojen neurologiset kliiniset oireet. J. Vet. Harjoittelija. Med. 31, 1209–1214. doi: 10.1111 / jvim.14728
PubMed Abstract | CrossRef Full Text | Google Scholar
Donato, C., and Vijaykrishna, D. (2017). Nisäkkäiden ja lintujen astrovirusten laaja isäntäalue ja geneettinen monimuotoisuus. Virukset 9: E102. doi: 10.3390 / v9050102
PubMed Abstract / CrossRef Full Text | Google Scholar
Drummond, A. J., Suchard, M. A., Xie, D., and Rambaut, A. (2012). Bayesilainen fylogenetiikka Beautin ja pedon kanssa 1.7. Mol. Biol. Evol. 29, 1969–1973. doi: 10.1093 / molbev/mss075
PubMed Abstract / CrossRef Full Text | Google Scholar
Giannitti, F., Anderson, M., Miller, M., Rowe, J., Sverlow, K., Vasquez, M., et al. (2016). Chlamydia pecorum: sikiön ja istukan leesiot satunnaisissa vuohien aborteissa. J. Vet. Diagn. Sijoittaa. 28, 184–189. doi: 10.1177/1040638715625729
PubMed Abstrakti / CrossRef kokoteksti / Google Scholar
Hirashima, Y., Okada, D., Shibata, S., Yoshida, S., Fujisono, S., Omatsu, T., et al. (2018). Koko genomin analyysi uudesta neurotrooppisesta naudan astroviruksesta, joka on havaittu japanilaisessa mustassa Härässä, jolla on ei-märkivä enkefalomyeliitti Japanissa. Kaari. Virol. 163, 2805–2810. doi: 10.1007 / s00705-018-3898-3
PubMed Abstrakti / CrossRef kokoteksti / Google Scholar
International Committee on Taxonomy of Viruses (2018). Master Species List (MSL32), Versio 1. Saatavilla: https://talk.ictvonline.org/files/master-species-lists/m/msl/7185 (accessed March, 2019).
Jenkins, G. M., Rambaut, A., Pybus, O. G., and Holmes, E. C. (2002). RNA-virusten molekyylievoluutiot: kvantitatiivinen fylogeneettinen analyysi. J. Mol. Evol. 54, 156–165. doi: 10.1007 / s00239-001-0064-3
PubMed Abstrakti / CrossRef kokoteksti / Google Scholar
Kumar, S., Stecher, G., and Tamura, K. (2016). MEGA7: molecular evolutionary genetics analysis version 7.0 isommille aineistoille. Mol. Biol. Evol. 33, 1870–1874. doi: 10.1093 / molbev/msw054
PubMed Abstract / CrossRef Full Text | Google Scholar
Li, L., Diab, S., McGraw, S., Barr, B., Traslavina, R., Higgins, R., et al. (2013). Poikkeava astrovirus, joka liittyy naudan neurologiseen sairauteen. Emerg. Tartuttaa. Tämä. 19, 1385–1392. doi: 10.3201 / eid1909.130682
PubMed Abstract | CrossRef Full Text | Google Scholar
Lole, K. S., Bollinger, R. C., Paranjape, R. S., Gadkari, D., Kulkarni, S. S., Novak, N. G., et al. (1999). Ihmisen immuunikatoviruksen tyypin 1 genomit, jotka on saatu C-alatyypin serokonvertereista Intiasta, ja intersubtype rekombinaatiosta. J. Virol. 73, 152–160.
PubMed Abstract / Google Scholar
Naccache, S. N., Peggs, K. S., Mattes, F. M., Phadke, R., Garson, J. A., Grant, P., et al. (2015). Neuroinvasiivisen astrovirusinfektion diagnoosi immuunipuutteisella aikuisella, jolla on enkefaliitti, puolueettoman seuraavan sukupolven sekvensoinnin avulla. Clin. Tartuttaa. Tämä. 60, 919–923. doi: 10.1093 / Cid/ciu912
PubMed Abstract | CrossRef Full Text | Google Scholar
Palmieri, C., Franca, M., Uzal, F., Anderson, M., Barr, B., Woods, L., et al. (2011). Psittaciformes-bakteerin Länsi-Niilin virustartunnan patologia ja immunohistokemialliset löydökset. Eläinlääkäri. Pathol. 48, 975–984. doi: 10.1177/0300985810391112
PubMed Abstract | CrossRef Full Text | Google Scholar
Pfaff, F., Schlottau, K., Scholes, S., Courtenay, A., Hoffmann, B., Höper, D., et al. (2017). Uusi astrovirus, joka liittyy enkefaliittiin ja ganglioneuriittiin kotimaisilla lampailla. Transbound. Emerg. Tämä. 64, 677–682. doi: 10.1111 / tbed.12623
PubMed Abstract | CrossRef Full Text | Google Scholar
Quan, P. L., Wagner, T. A., Briese, T., Torgerson, T. R., Hornig, M., Tashmukhamedova, A., et al. (2010). Astrovirus-enkefaliitti pojalla, jolla on X-linkitetty agammaglobulinemia. Emerg. Tartuttaa. Tämä. 16, 918–925. doi: 10.3201 / eid1606. 091536
PubMed Abstract | CrossRef Full Text | Google Scholar
Rambaut, A., Lam, T. T., Max Carvalho, L., and Pybus, O. G. (2016). Tutkitaan heterokronisten sekvenssien ajallista rakennetta Tempestin (formely Path-O-Gen) avulla. Virus Evol. 2: vew007. doi: 10.1093 / ve / vew007
PubMed Abstract / CrossRef Full Text | Google Scholar
Reuter, G., Pankovics, P., and Boros, A. (2018). Ei-suppuratiivinen (aseptinen) meningoenkefalomyeliitti, joka liittyy neurovirulenttisiin astrovirusinfektioihin ihmisillä ja eläimillä. Clin. Mikrobiolia. Rev. 31, e00040-18. doi: 10.1128 / CMR.00040-18
PubMed Abstract | CrossRef Full Text | Google Scholar
Schlottau, K., Schulze, C., Bilk, S., Hanke, D., Höper, D., Beer, M., et al. (2016). Uuden naudan astroviruksen havaitseminen lehmässä, jolla on enkefaliitti. Transbound. Emerg. Tämä. 63, 253–259. doi: 10.1111 / tbed.12493
PubMed Abstract | CrossRef Full Text | Google Scholar
Selimovic-Hamza, S., Boujon, C. L., Hilbe, M., Oevermann, A., and Seuberlich, T. (2017a). Bovine astrovirus CH13/NeuroS1 infection in neurologically-disease cattle: towards assessment of causality. Virukset 9: E12. doi: 10.3390 / v9010012
PubMed Abstract / CrossRef Full Text | Google Scholar
Selimovic-Hamza, S., Sanchez, S., Philibert, H., Clark, E. G., and Seuberlich, T. (2017b). Bovine astrovirus infektio rehukarjan neurologinen sairaus Länsi-Kanadassa. Voi. Eläinlääkäri. J. 58, 601-603.
Google Scholar
Selimovic-Hamza, S., Bouzalas, I. G., Vandevelde, M., Oevermann, A., and Seuberlich, T. (2016). Astroviruksen osoittaminen Euroopan sporadisen naudan enkefaliitin historiallisissa tapauksissa Sveitsissä 1958-1976. Edessä. Eläinlääkäri. Sci. 3:91. doi: 10.3389 / fvets.2016.00091
PubMed Abstract | CrossRef Full Text | Google Scholar
Seuberlich, T., Wüthrich, D., Selimovic-Hamza, S., Drögemüller, C., Oevermann, A., Bruggmann, R., et al. (2016). Toisen enkefaliittiin liittyvän astroviruksen tunnistaminen naudoissa. Emerg. Mikrobit Tarttuvat. 5: e5. doi: 10.1038 / emi.2016.5
PubMed Abstract | CrossRef Full Text | Google Scholar
Spinato, M. T., Vince, A., Cai, H., and Ojkic, D. (2017). Naudan astroviruksen tunnistaminen naudan muuta kuin märkivää enkefaliittia sairastavissa tapauksissa Itä-Kanadassa. Voi. Eläinlääkäri. J. 58, 607-609.
Google Scholar
Stein, L. T., Rech, R. R., Harrison, L., and Brown, C. C. (2010). Immunohistokemiallinen tutkimus rabiesviruksesta koti-ja villieläinlajien keskushermostossa. Eläinlääkäri. Pathol. 47, 630–633. doi: 10.1177/0300985810370013
PubMed Abstrakti / CrossRef kokoteksti / Google Scholar
Trifinopoulos, J., Nguyen, L. T., von Haeseler, A., and Minh, B. Q. (2016). W-IQ-TREE: nopea online-fylogeneettinen työkalu maksimaaliseen todennäköisyysanalyysiin. Nukleiinihapot Res. 44, W232–W235. doi: 10.1093 / nar/gkw256
PubMed Abstract / CrossRef Full Text | Google Scholar
Tse, H., Chan, W. M., Tsoi, H. W., Fan, R. Y., Lau, C. C., Lau, S. K., et al. (2011). Naudan astrovirusten uudelleen löytäminen ja genomien luonnehtiminen. J. Kenr. Virol. 92 (Pt 8), 1888-1998. doi: 10.1099 / vir.0, 030817-0
PubMed Abstract | CrossRef Full Text / Google Scholar