Solubiologia 06: the Cytoskeleton Part II: Tubulin
These are notes from lecture 6 of Harvard Extension ’ s Cell Biology course.
viime viikolla käsittelimme aktiinista valmistettuja mikrofilamentteja. Tällä viikolla: tubuliinista valmistetut Mikrotubulukset. Miksi tukiranka tarvitsee kaksi erillistä järjestelmää? Ajattele, että mikrofilamentit ovat vain yksittäisten alayksiköiden ketjuja, kun taas Mikrotubulukset ovat kirjaimellisesti putkia (kuten pian näemme) – onttoja sylintereitä, joiden seinät on tehty dimeereistä tehdyistä ketjuista. Nämä erilaiset rakenteet tarkoittavat erilaisia ominaisuuksia: mikrofilamentit voivat helpommin muodostua spontaanisti, ja ne voivat haarautua (Arp2/3: n ja muiden Viimeksi käsiteltyjen kompleksien avulla) muodostaakseen erilaisia verkkomuotoja ja yhdistääkseen solun eri osia. Mikrotubulukset tukeutuvat voimakkaammin nukleaatioon muodostuakseen, ja ovat pohjimmiltaan puheverkko, joka yhdistää centrosomin solun reuna-alueisiin. Käytännössä mikrofilamentteja on solukuoressa runsaasti ja ne osallistuvat voimakkaasti supistumiskykyisiin liikkeisiin ja solujen liikkuvuuteen, kun taas Mikrotubulukset osallistuvat eniten organellien järjestämiseen ja toimivat ratoina anterogradi-ja retrogradiseen kuljetukseen.
Mikrotubulukset
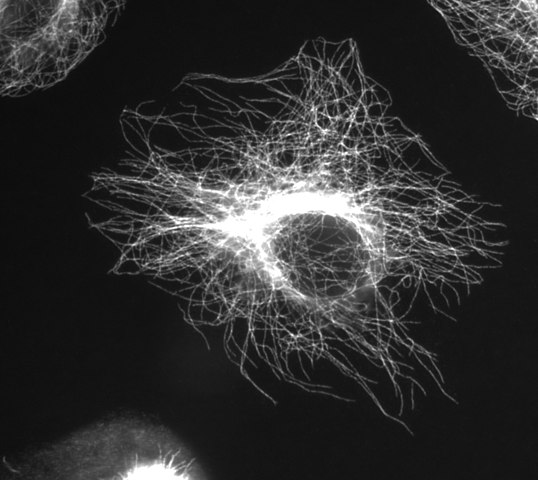
(kuva kiitos Wikimedia Commons-käyttäjä Jeffrey81)
Wikipedian lisäksi NCBI: llä saatavilla olevien lodishin ja Cooperin solubiologian oppikirjojen aiemmat painokset ovat myös erinomaisia referenssejä.
mikrotubulusten putkimainen rakenne on tukevampi kuin mikrotubulusten, mikä mahdollistaa paljon raskaan vetämisen ja työntämisen. Vaikka Mikrotubulukset ovat lujia, ne ovat lämpötilasta riippuvaisia-ne depolymeroituvat, jos ne jäähdytetään 4°C: seen, ja repolymeroituvat uudelleen, jos ne kuumennetaan 37°C: seen edellyttäen, että GTP on käytettävissä.
mikrotubulusten perusrakenneosa on α-tubuiliini/β-tubuliinidimeeri (jota koodaavat TUBA_-geenit ja TUBB_-geenit). Yksittäiset tubuliiniproteiinit painavat kukin ~55kda, joten ~110da / aminohappo, joka on 500 aminohapon luokkaa. Sekä α-että β-tubuliini sitovat GTP: tä, mutta α melko pitkälti vain pitää siitä kiinni ikuisesti, kun taas β: ssa se voidaan vaihtaa tai hydrolysoida BKT: ksi ja vaihtaa sitten taas uudeksi GTP: ksi.
peräkkäisten α/β-dimeerien säikeet muodostavat protofilamentteja, ja 13 sylinteriin vierekkäin asetettua protofilamenttia muodostavat mikrotubulin. Protofilamenttien välissä olevaa tilaa kutsutaan ”saumaksi”. Koko mikrotubulin halkaisija on ~25 nm. Voit tavallaan nähdä sen rakenteen (joka on valmistettu dimeereistä ja protofilamenteista) mikrotubulusten Solusegmentin sisäisessä elämässä.:
kunkin dimeerin sisällä β – alayksikkö on polymeroitumisessa suosittu (+) pää ja α on depolymeroitumisessa suosittu (-) Pää. Mikrotubulukset muodostavat periaatteessa samat kolme vaihetta kuin mikrofilamentit: nukleaation, venymän ja vakaan tilan. Mutta toisin kuin mikrofilamentit ne eivät nukleoidu helposti itsestään, joten nukleaatio vaatii mikrotubulusten järjestelykeskuksia (MTOCs). Jakautumattomissa soluissa on kussakin vain yksi MTOC, jota kutsutaan centrosomiksi. (Tämä on kuvattu myös yllä olevalla videolla). Sijaitsee lähellä tumaa, centrosome on napa säteittäinen konfiguraatio Mikrotubulukset ( – ) päät osoittavat sisään ja ( + ) päät osoittavat solun reuna.
sentrosomi koostuu 2 sylinteristä, joita kutsutaan sentrioleiksi. Jokainen sentrioli koostuu jokaisesta 9 sarjasta 3 lateraalisesti fuusioitunutta mikrotubulusta, joita ympäröi amorfinen perikentriolaarinen materiaali, joka sisältää runsaasti nukleaatiota edistäviä asioita-erityisesti γ-tubuliinirengaskomplekseja (gammatubuliinia koodaavat TUBG_-geenit). γ-tubuliinia pidetään ”halkaistuna pesukoneena”:

mallin mukaan halkaistujen päiden avulla γ-tubuliini sitoutuu α-tubuliiniin eli muodostuvan mikrotubuluksen ( – ) päähän, jolloin muodostuu nukleaatiosiemen mikrotubuluksen muodostumiselle.
Mikrotubuluksia voi muodostua in vitro. Dynamiikka riippuu enimmäkseen kriittisistä pitoisuuksista. (- )- Pää on vähemmän polymeroitumiseen taipuvainen kuin ( + ) – pää, joten sillä on suurempi kriittinen pitoisuus. Jos todellinen pitoisuus on kahden pään kriittisten pitoisuuksien välillä, juokseminen tapahtuu.
kun mikrotubulus yhtäkkiä alkaa hajota, tätä kutsutaan ”katastrofiksi”. Katastrofeissa on mielenkiintoista energistä dynamiikkaa. Muistuttaa aiemmin, että beetatubuliini, joka on ( + ) pää, jossa venymä tapahtuu, voi olla joko GTP – tai BKT-sidottu. Se on vakaampi osa mikrotubulustaan, kun se on GTP: hen sidottu; BKT: hen sidottu tubuliini on altis dissosioitumaan. Mikrotubulukset muodostuvat pääasiassa lisäämällä GTP: hen sitoutunutta beetatubuliinia (+) päähän, mutta lisättyään beetatubuliinimolekyylit hydrolysoivat myöhemmin GTP: nsä, jolloin ne jäävät BKT: hen sitoutuneiksi. Joten on olemassa eräänlainen ”kärki” mikrotubulus, joka on GTP-sidottu, kun taas beetatubuliini syvemmälle mikrotubulus, lisätty pidempään sitten, ovat BKT-sidottu. Jos GTP: n hydrolyysin raja saavuttaa kärjen tai jos mikrotubulus katkaistaan, vähemmän stabiili, BKT: hen sidottu beetatubuliini paljastuu ja protofilamentit alkavat irrota kuin ”Ramin sarvet”. Jos näin tapahtuu, mikrotubuli hajoaa, kunnes se osuu GTP: hen sitoutuneen beetatubuliinin ”saarekkeeseen” jossain kauempana oljesta. Katastrofi voidaan ’pelastaa’ lisäämällä uusia GTP-sidottuja tubuliinidimenttejä, jotka peittävät päät ja vakauttavat protofilamentit, jolloin mikrotubuli voi muodostua uudelleen. Näet tästä videosta, että mikrotubulusten depolymerointi voi olla paljon nopeampaa kuin polymerointi:
tässä on joitakin mikrotubulusten tutkimiseen hyödyllisiä yhdisteitä. Kihtilääke kolkisiini sitoo vapaita alfa-beeta-dimeerejä, mikä vähentää niiden tarjontaa mikrotubulusten muodostumisessa ja edistää siten depolymerisaatiota. Nokodatsoli häiritsee myös uusien mikrotubulusten muodostumista, ja koska uusien mikrotubulusten muodostuminen on tärkeää mitoosin kannalta, nokodatsoli on Antineoplastinen syöpälääke. Toisaalta paklitakseli (taxol), toinen syöpälääke, toimii stabiloimalla mikrotubuluksia, koska olemassa olevien mikrotubulusten hajoaminen on tärkeää myös mitoosin kannalta.
Mikrotubulukseen liittyvillä proteiineilla (MAPs) on erilaisia rooleja. Huomionarvoisia ovat MAP4 (ei-hermosoluissa), MAP2 (neuroneissa) ja Tau (hermosoluissa; koodattu MAPT-geenillä). Kaikille näille on yhteistä, että ne toimivat mikrotubulusten vakauttamiseksi, siirtäen kinetiikkaa mikrotubulusten kasvun hyväksi ja katastrofia vastaan. Jokaisessa on 18 aminohappojousitus, jossa on positiivisesti varautuneita aminohappoja, jotka sitoutuvat mikrotubulusten negatiiviseen osaan. Ne saattavat niputtaa useita mikrotubuluksia yhteen , jolloin MAP2 pitää Mikrotubulukset pidemmällä etäisyydellä, koska niiden varsi on pidempi (Tau: iin verrattuna).
karttoja säännellään fosforyloinnilla: MARK-proteiinikinaasit sitovat kovalenttisesti fosfaattiryhmiä S -, T-tai Y-aminohapoiksi MAP-proteiineissa, mikä vähentää niiden kykyä sitoa mikrotubuluksia. (Sykliiniriippuvaiset kinaasit säätelevät karttoja myös mitoosin aikana). Näiden karttojen arvellaan määrittävän solujen fysikaalisen rakenteen. Neuroneissa MAP2 on dendriiteissä ja Tau on enimmäkseen aksonissa. Tau: ta koodaavan MAPT-geenin mutaatiot aiheuttavat frontotemporaalista dementiaa (FTD). Hyperfosforyloitua Tau: ta löytyy (vaikka emme tiedä miksi) Alzheimer-potilaiden aivoista. Molempien sairauksien hiirimalleissa näkyy aksonaalinen degeneraatio, mikä sopii siihen, että Tau ei pysty tekemään työtään aksonin mikrotubulusten vakauttamiseksi.
toinen proteiiniluokka, jonka nimi on +kärjet niiden sitoutumisesta mikrotubulusten (+) päähän, voi suojata katastrofilta. Ne näyttävät ulottuvan reilusti mikrotubulukseen. Videolla EB-1 työntyy ulospäin, koska mikrotubulusten päät kasvavat:
muut päätettä sitovat proteiinit edistävät katastrofia stabiilisuuden sijaan. Kinesin-13 toimii protofilamenttien lopun käyristämiseksi alentaen katastrofin kynnystä. Stathmin (STMN1 geeni; aka Op18, jossa Op tarkoittaa onkoproteiinia) sitoutuu tubuliinidimeereihin protofilamentin sisällä, edistäen sekä välitöntä katastrofia että mahdollisesti myös GTP-hydrolyysiä, joka poistamalla ”saaret” GTP-sitoutuneesta beetatubuliinista edustaisi enemmän pitkän aikavälin investointeja katastrofin hyväksi. Stathmiinia säädellään fosforylaatiolla. Katanin (japanilaisen Katana-miekan mukaan) katkaisee kirjaimellisesti Mikrotubulukset.
Mikrotubulusmotoriset proteiinit ovat pitkälti samanlaisia kuin mikrotubulusmotoriset proteiinit. Niitä on kaksi sukua: kinesiinit, joista suurin osa liikkuu anterogradilla eli kohti ( + ) päätä, ja dyneiinit, joista suurin osa liikkuu retrogradisesti eli kohti ( – ) päätä. Ne ”kävelevät” näin:
Kinesiini ja dyneiini osallistuvat organellien liikkumiseen, endosytoosiin ja eksosytoosiin sekä kromosomien eriytymiseen meioosin aikana & mitoosi.
Kinesiineistä parhaiten tutkittu Kinesiini-1 on ”tavanomainen” kinesiini siinä mielessä, että se toimii kahdesta raskaasta ketjuyksiköstä (KIF1-geenit esim. KIF1A), jotka koostuvat päädomeeneista, jotka hydrolysoivat ATP: tä ja sitovat mikrotubuluksia, ja kahdesta kevyestä ketjuyksiköstä (KLC-geenit, esim.KLC1), jotka muodostavat ”hännän”, joka sitoo lastin, tässä tapauksessa vesikkelit. Linker domain (mikä proteiini on tämä osa?) mahdollistaa kahden raskaan ketjun dimerisoinnin.
tässä video siitä toiminnassa. Huomaa, että video kutsuu sitä dimeeri sijasta tetrameeri; luulen, että se johtuu siitä, että he vain harkitsee päät eikä keskustella hännät.
jotkin muut kinesiinit eroavat hieman toisistaan:
- Kinesiini-2, joka tekee myös vesikkelin & organellenkuljetuksen, on heterotrimeeri, jolla on kaksi erilaista (joskin toisiinsa liittyvää) raskasta ketjua ja toinen polypeptidi, jota se käyttää lastinsa säätelyyn.
- Kinesiini-5: ssä on pään ja hännän sijaan päät molemmissa päissä, joten se kulkee vastakkaisiin suuntiin kahdella eri mikrotubuluksella vetäen ne yhteen. Tätä kutsutaan bipolaariseksi liikkeeksi.
- Kinesiini-14 on ainoa tunnettu kinesiini, joka liikkuu kohti (-) loppua, ja se osallistuu mitoosiin.
”kävelyliikkeen” periaatteet ovat kuitenkin jokseenkin samat kaikille kinesiineille, ja ne on kuvattu yllä olevalla viimeisellä videolla. Kun ”päät” eivät ole sitoutuneet mikrotubulukseen, ne ovat molemmat ADP-sidottuja. Yksi pää sattuu kohtaamaan mikrotubulin ja sitoutuu siihen vapauttaen sen ADP: n, jolloin ATP korvaa sen. ATP-sidos indusoi konformaatiomuutoksen, joka vetää linkkeriä, heiluttaa toista, ”laahaa” päätä eteenpäin johtavaan asentoon, jossa se sitoutuu mikrotubulukseen. Tämän jälkeen alkuperäinen Pää hydrolysoi ATP: n, joka vapauttaa fosfaattia (joka on videossa lyhennetty PI: ksi) ja antaa energiaa tämän prosessin yhteen energeettiseen ylämäkeen, joka irtoaa mikrotubuluksesta.
Myosiinilla (mikrofilamenteilla kulkeva motorinen proteiini) ja kinesiinillä on hyvin samanlaiset rakenteet, mutta ei aminohappojärjestyksen samankaltaisuutta. Niiden ei siis uskota olevan lamaannuttavia, vaan pikemminkin esimerkki konvergentista evoluutiosta.
ihmiset puhuvat paljon kinesiineistä osittain siksi, että emme ymmärrä dyneinejä aivan yhtä hyvin. Dyneiinit ovat valtavia proteiineja, jotka koostuvat 2 suuresta, 2 väli-ja 2 pienestä alayksiköstä. Niiden pelkkä koko on tehnyt niistä vaikeasti eristettäviä ja luonnehdittavia, eikä niiden toimintaa tunneta kovin hyvin. Tiedämme, että dynactin-kompleksi (moniproteiinikompleksi; dynactiinit itse ovat dtcn_-geenejä) on mukana ”adapterina”, joka yhdistää dyneinin lastiin.
tämä video tiivistää koko tukirangan ja köydet suureen osaan viime viikon ja tämän luennon aineistosta:
PrP
kuten eritysreitin muistiinpanoissa mainitaan, PRP on GPI-ankkuroitunut kalvoon ja käy läpi endosytoosin hyvin säännöllisesti, jolloin syntyy endosyyttisiä vesikkeleitä, joissa on PrP: tä. Encalada 2011 toteaa, että Kinesin-1c ja DHC1 (dynein heavy chain 1) ovat vastuussa näiden PrP-vesikkelien anterogradisesta ja retrogradisesta kuljettamisesta. Kun Encalada tyrmäsi, kaatoi tai esti Kinesiini-1c: tä, anterogradi ja retrogradinen liike vähenivät, ja vastaavasti DHC1: n toiminta häiriintyi. Näyttää siis siltä, että nämä kaksi kompleksia, vaikka ne liikkuvat vastakkaisiin suuntiin, aktivoivat toisiaan. Ja mielenkiintoista on, että näiden motoristen proteiinien häiritseminen ei estänyt PRP – vesikkeleitä liittymästä moottoreihin-ne eivät vain liikkuneet yhtä nopeasti tai yhtä usein. Paperissa on useita muita solubiologian johtopäätöksiä siitä, miten vesikkelit aktivoivat motorisia proteiineja ja miten kuljetussuunta määritetään.