Sejtbiológia 06: a citoszkeleton II. rész: Tubulin
ezek a Jegyzetek a Harvard Extension Sejtbiológiai tanfolyamának 6.előadásából.
a múlt héten aktinból készült mikrofilamentumokat fedtünk le. Ezen a héten: tubulinból készült mikrotubulusok. Miért, kérdezed, a citoszkeletonnak két külön rendszerre van szüksége? Vegyük figyelembe, hogy a mikrofilamentumok csak az egyes alegységek láncai, míg a mikrotubulusok szó szerint csövek (Amint hamarosan látni fogjuk) – üreges hengerek, amelyek falai dimerekből készült láncokból készülnek. Ezek a különböző struktúrák különböző képességeket jelentenek: a mikrofilamentumok könnyebben spontán módon alakulhatnak ki, és elágazhatnak (az Arp2/3 és más, Legutóbb tárgyalt komplexek segítségével) annak érdekében, hogy különböző hálózati alakzatokat alakítsanak ki és összekapcsolják a sejt különböző részeit. A mikrotubulusok nagyobb mértékben támaszkodnak a magképződésre annak érdekében, hogy kialakuljanak, és alapvetően egy küllős hálózat, amely összeköti a centroszómát a sejt perifériájával. A gyakorlatban a mikroszálak bőségesek a sejtkéregben, és erősen részt vesznek a kontraktilis mozgásokban és a sejtek motilitásában, míg a mikrotubulusok leginkább az organellák szervezésében és az anterográd és retrográd transzport nyomvonalaként szolgálnak.
mikrotubulusok
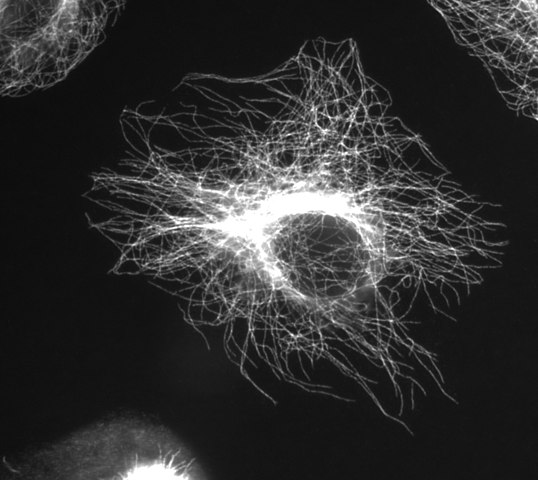
(Image thanks to Wikimedia Commons user Jeffrey81)
a Wikipédián kívül az NCBI-n elérhető Lodish és Cooper cell biology tankönyvek korábbi kiadásai is kiváló referenciák.
a mikrotubulusok csőszerkezete szilárdabb, mint a mikrofilamentumok, így sok nagy teherbírású húzást és tolást tesz lehetővé. Bár erős, a mikrotubulusok hőmérséklet-függőek-depolimerizálódnak, ha lehűtik 4 6CC-ra, és újra repolimerizálódnak, ha 37 C-ra melegítik, feltéve, hogy GTP áll rendelkezésre.
a mikrotubulusok alapvető építőeleme egy (a TUBA_ gének, illetve a TUBB_ gének által kódolt) ~ -tubuilin/ ~ -tubulin dimer. Az egyes tubulinfehérjék súlya ~55kda, tehát ~110da / aminosav esetén ez 500 aminosav nagyságrendű. Mind a~, mind a ~ ~ tubulin megköti a GTP-t, de a ~ nagyjából csak örökké tartja, míg a ~ ~ ban cserélhető vagy hidrolizálható a GDP-re, majd újra kicserélhető új GTP-re.
egymást követő, egymást követő, egymást követő, egymást követő, egymást követő, egyenként hengerbe rendezett, 13 darab protofilamentum alkot egy mikrotubulust. A protofilamentumok közötti teret varratnak nevezzük. A teljes mikrotubulus átmérője ~25 nm. Látható a szerkezete (dimerekből és protofilamentumokból) a Sejtszegmens belső életében a mikrotubulusokról:
az egyes dimereken belül a (+) vég a polimerizáció szempontjából előnyben részesített ( + ) vég, a ( – ) vég pedig a depolimerizáció szempontjából kedvelt. A mikrotubulusok alapvetően ugyanabban a három lépésben alakulnak ki, mint a mikrofilamentumok: magképződés, nyúlás és egyensúlyi állapot. De a mikrofilamentumokkal ellentétben önmagukban nem nukleálódnak könnyen, ezért a nukleációhoz mikrotubulus-szervező központokra (Mtoc) van szükség. A nem osztódó sejteknek csak egy MTOC – ja van, az úgynevezett centroszóma. (Ezt a fenti videó is ábrázolja). A mag közelében elhelyezkedő centroszóma a mikrotubulusok radiális konfigurációjának központja, amelynek ( – ) végei a sejt perifériájára mutatnak, a ( + ) végei pedig a sejt perifériájára mutatnak.
a centroszóma 2 hengerből áll, amelyeket centrioloknak neveznek. Minden centriol mindegyik 9 db 3 oldalirányban fuzionált mikrotubulusból áll, amorf ‘pericentrioláris anyag’ veszi körül, amely gazdag olyan dolgokban, amelyek elősegítik a nukleációt – különösen az Alzheimer-tubulin gyűrűs komplexeket (a gamma-tubulint a TUBG_ gének kódolják). a tubulint úgy tekintik, mint egy osztott alátétet:

a modell az, hogy a hasított végek lehetővé teszik a vállalkozók számára, hogy a (-) tubulin kötődjön a (-) tubulinhoz, vagyis a ( – ) végéhez, amely egy mikrotubulus kialakulásához szükséges magképződést biztosít.
mikrotubulusok képződhetnek in vitro. A dinamika leginkább a kritikus koncentrációktól függ. A ( – ) vég kevésbé hajlik a polimerizáció felé, mint a ( + ) vég, ezért nagyobb a kritikus koncentrációja. Ha a tényleges koncentráció a két vég kritikus koncentrációja között van, futópadozás történik.
amikor egy mikrotubulus hirtelen disszociálni kezd, ezt katasztrófának nevezzük. A katasztrófáknak érdekes energetikai dinamikájuk van. Emlékezzünk vissza korábban, hogy a béta-tubulin, amely a ( + ) vég, ahol a megnyúlás történik, lehet GTP – vagy GDP-kötött. Mikrotubulusának stabilabb része, ha GTP-hez kötött; a GDP-hez kötött tubulin alkalmas disszociálni. A mikrotubulusok elsősorban GTP-hez kötött béta-tubulin hozzáadásával alakulnak ki a ( + ) végén, de hozzáadásuk után a béta-tubulin molekulák később hidrolizálják GTP-jüket, így GDP-kötődnek. Tehát van egyfajta ‘csúcs’ a mikrotubulusnak, amely GTP-kötött, míg a béta-tubulin mélyebben a mikrotubulusban, hosszabb ideig hozzáadva, GDP-kötött. Ha a GTP hidrolízis határa eléri a csúcsot, vagy ha valami történik a mikrotubulus levágásával, akkor a kevésbé stabil, GDP-hez kötött béta-tubulin ki van téve, és a protofilamentumok elkezdenek hámozni, mint a “Kos szarvai”. Ha ez megtörténik, a mikrotubulus szétesik, amíg el nem éri a GTP-hez kötött béta-tubulin “szigetét” valahol a szalma alatt. A katasztrófa megmenthető új GTP-vel kötött tubulin dimerek hozzáadásával, amelyek lezárják a végeket és stabilizálják a protofilamentumokat, lehetővé téve a mikrotubulus újbóli kialakulását. Ebben a videóban láthatja, hogy a mikrotubulusok depolimerizációja sokkal gyorsabb lehet, mint a polimerizáció:
Íme néhány vegyület, amely hasznos a mikrotubulusok tanulmányozásához. A kolhicin, egy köszvényellenes gyógyszer, megköti a szabad alfa-béta dimereket, csökkentve a mikrotubulusok képződésének ellátását, ezáltal elősegítve a depolimerizációt. A nocodazol az új mikrotubulusok kialakulását is befolyásolja, és mivel az új mikrotubulusok kialakulása fontos a mitózis szempontjából, a nocodazol daganatellenes rákellenes gyógyszer. Ezzel szemben a paklitaxel (taxol), egy másik rákellenes gyógyszer, a mikrotubulusok stabilizálásával működik, mivel a meglévő mikrotubulusok lebontása szintén fontos a mitózis szempontjából.
a Mikrotubulushoz kapcsolódó fehérjéknek (MAPs) különböző szerepük van. Figyelemre méltó a MAP4 (nem neuronális sejtekben), a MAP2 (neuronokban) és a tau (neuronokban; a mapt gén kódolja). Mindezekben közös, hogy stabilizálják a mikrotubulusokat, a kinetikát a mikrotubulusok növekedésének javára és a katasztrófa ellen változtatják. Mindegyik tartalmaz egy 18 aminosav-nyújtást pozitív töltésű aminosavakkal, amelyek kötődnek a mikrotubulusok negatív részéhez. Több mikrotubulust köthetnek össze, a MAP2 hosszabb karja miatt nagyobb távolságban tartja a mikrotubulusokat (a Tau-hoz képest).
a térképeket foszforilezéssel szabályozzák: a MARK protein kinázok kovalensen kötik a foszfátcsoportokat a map fehérjék S, T vagy Y aminosavaihoz, csökkentve a mikrotubulusok kötődésének képességét. (A ciklinfüggő kinázok a mitózis során is szabályozzák a térképeket). Úgy gondolják, hogy ezek a térképek meghatározzák a sejtek fizikai szerkezetét. Az idegsejtekben a MAP2 a dendritekben található, a Tau pedig többnyire az axonban található. A Tau-t kódoló mapt gén mutációi frontotemporális demenciát (FTD) okoznak. A hiperfoszforilált Tau megtalálható (bár nem tudjuk, miért) az Alzheimer-kórban szenvedő betegek agyában. Mindkét betegség egérmodelljei axonális degenerációt mutatnak, összhangban azzal, hogy a Tau nem képes elvégezni a mikrotubulusok stabilizálását az axonban.
a fehérjék egy másik osztálya, a +tippek a mikrotubulusok (+) végéhez való kötődésükhöz, védelmet nyújt a katasztrófa ellen. Úgy tűnik, hogy kiterjeszti a tisztességes módon le a mikrotubulus. Ez a videó azt mutatja, hogy az EB-1 kifelé tolódik, mert a mikrotubulusok végei nőnek:
más végkötő fehérjék elősegítik a katasztrófát, nem pedig a stabilitást. A kinezin-13 a protofilamentumok végének görbületére hat, csökkentve a katasztrófa küszöbét. Stathmin (STMN1 gén; más néven Op18, ahol az Op az onkoproteint jelenti) kötődik a tubulin dimerekhez egy protofilen belül, elősegítve mind az azonnali katasztrófát, mind esetleg a GTP-hidrolízist, amely a GTP-hez kötött béta-tubulin “szigeteinek” eltávolításával inkább hosszú távú befektetést jelentene a katasztrófa javára. A Stathmin foszforilezéssel van szabályozva. Katanin (a japán kard katana után) szó szerint elvágja a mikrotubulusokat.
a mikrotubulus motoros fehérjék nagyrészt hasonlóak a mikrofilament motoros fehérjékhez. Két családba tartoznak: kinezinek, amelyek többsége anterográdot, azaz a (+) vég felé mozog; és dyneinek, amelyek többsége retrográd, azaz a (-) vég felé mozog. Így járnak:
a kinezin és a dynein részt vesz a mozgó organellákban, az endocitózisban és az exocitózisban, valamint a kromoszóma szegregációban a meiózis során & mitózis.
a kinezinek közül a legjobban tanulmányozott kinezin-1 egy hagyományos kinezin, mivel tetramer-ként működik, amely két nehéz láncegységből áll (KIF1 gének pl. KIF1A), amelyek az ATP hidrolizálására és a mikrotubulusok megkötésére szolgáló ‘fej’ doméneket, valamint két könnyű lánc egységet (KLC gének, pl. KLC1) tartalmaznak, amelyek a rakományt megkötő ‘farokot’, ebben az esetben vezikulumokat tartalmaznak. Linker domén (melyik fehérje ez a része?) lehetővé teszi két nehéz lánc dimerizálását.
itt van egy videó róla működés közben. Ne feledje, hogy a videó dimernek hívja tetramer helyett; azt hiszem, ez azért van, mert csak a fejeket veszik figyelembe, a farokról nem beszélnek.
néhány más kinezin kissé különbözik:
- a kinezin-2, amely a & organellát is szállítja, heterotrimer, két különböző (bár rokon) nehéz lánccal és egy másik polipeptiddel, amelyet rakományának szabályozására használ.
- a kinezin-5-nek mindkét végén feje van a fej és a farok helyett, így két különböző mikrotubuluson ellentétes irányban halad, összehúzza őket. Ezt bipoláris mozgásnak nevezik.
- a kinezin-14 az egyetlen ismert kinezin, amely a (-) vég felé mozog, és részt vesz a mitózisban.
a ‘walking’ mozgalom alapelvei nagyjából megegyeznek az összes kinezin esetében, és a fenti videóban is ez látható. Ha nem kötődnek mikrotubulushoz, akkor a’ fejek ‘ mind ADP-kötöttek. Az egyik fej véletlenül találkozik egy mikrotubulussal, és kötődik hozzá, felszabadítva az ADP-jét, lehetővé téve az ATP helyettesítését. Az ATP-kötés konformációs változást idéz elő, amely meghúzza a kapcsolót, a másikat lengve, a fejét előre haladva vezető helyzetbe, ahol a mikrotubulushoz kötődik. Az eredeti fej ezután hidrolizálja az ATP-t, amely felszabadítja a foszfátot (amelyet a videóban Pi-nek rövidítenek), és biztosítja az energiát a folyamat egy energetikailag felfelé vezető lépéséhez, amely elszakad a mikrotubulustól.
a miozin (a mikrofilamentumokon járó motoros fehérje) és a kinezin szerkezete nagyon hasonló, de nincs aminosav-szekvenciája. Így úgy gondolják, hogy nem paralogok, hanem inkább a konvergens evolúció példája.
az emberek sokat beszélnek a kinezinekről, részben azért, mert nem értjük a dyneineket is. A dyneinek hatalmas fehérjék, amelyek 2 nagy, 2 közbenső és 2 kis alegységből állnak. Puszta méretük miatt nehéz őket elkülöníteni és jellemezni, működésük pedig nem jól ismert. Tudjuk, hogy a dynactin komplex (többfehérje komplex; maguk a dynactinok a DTCN_ gének) egy adapterként vesz részt, amely összeköti a dyneint a cargo-val.
ez a videó összefoglalja az egész citoszkeletont és a köteleket a múlt hét anyagának nagy részében és ebben az előadásban:
PrP
amint azt a szekréciós útvonal megjegyzi, a PrP GPI-vel van rögzítve a membránhoz, és nagyon rendszeresen endocitózison megy keresztül, endocita vezikulákat hozva létre PrP-vel. Az Encalada 2011 megállapítja, hogy a kinezin-1c és a DHC1 (dynein heavy chain 1) felelősek ezen PrP vezikulák anterográd és retrográd szállításáért. Amikor az Encalada kiütötte, leütötte vagy gátolta a kinezin-1C-t, az anterográd és a retrográd mozgás csökkent, és hasonlóan, amikor a DHC1 megszakadt. Tehát úgy tűnik, hogy ez a két komplex, bár ellentétes irányban mozognak, aktiválják egymást. Érdekes módon ezeknek a motorfehérjéknek a megzavarása nem akadályozta meg a PrP vezikulák motorokhoz való kapcsolódását – csak nem mozogtak olyan gyorsan vagy olyan gyakran. A tanulmány számos más sejtbiológiai következtetést tartalmaz arról, hogy a vezikulák hogyan aktiválják a motoros fehérjéket, és hogyan határozzák meg a szállítás irányát.