微生物学のFrontiers
Introduction
アストロウイルス科には、哺乳動物と鳥類に感染するママストロウイルスとアバストロウイルスの二つの属に非エンベロープ、陽性センス、一本鎖RNAウイルスが含まれている。 現在、ウイルスの分類に関する国際委員会(International Committee on Taxonomy of Viruses,2018)は、ママストロウイルス属内の19種、すなわちママストロウイルス-1から-19を認識していますが、分類を待っている多くの株があり、そのうちのいくつかは暫定的に新種と考えられています(Donato and Vijaykrishna,2017)。
2010年以降、いくつかのアストロウイルスは、ヒトを含む様々な哺乳動物種において神経侵襲性としてますます認識されている(Quan et al. ら、2 0 1 0;Naccache e t a l. ら、2015)、ミンク(Blomströmら、2015)、ミンク(Blomström et al. ら、2 0 1 0)、ウシ(Li e t a l. ら、2 0 1 3)、羊(Pfaff e t a l. ら、2 0 1 7)、およびブタ(Borosら、2 0 1 8)、およびブタ(2 0 1 9)。, 2017). 米国牛におけるウシアストロウイルス関連脳炎の最初の認識後(Li et al. ら、2013)、スイスからの病因不明の散発性ウシ脳炎の症例における遡及的研究は、この神経侵襲性アストロウイルスが何十年も検出されなかったことを明, 2016). これらのアストロウイルスの疫学および伝達経路は不明であるが、ヌクレオチドおよびアミノ酸レベルで牛および卵の神経侵襲的なアストロウイルスの間で共有される高いレベルの同一性(>98%)に基づいて、種間伝達が示唆されている(Boujon et al., 2017).
ウシのアストロウイルス(Boastv)、BoAstV-Neurose1と命名された(Li et al. ら、2 0 1 3)およびBoastv−C H1 3(Bouzalasら、2 0 1 3)。,2014),最初に非化膿性脳炎の牛の脳で発見されました。,米国とスイスで,それぞれ. 異なる命名法にもかかわらず、両方のウイルスは同じ遺伝子型種を表す(Bouzalas e t a l. ら、2 0 1 6;Selimovic−Hamza e t a l. 2017a)は、ICTVによる公式の分類をまだ待っている。 2015年には、スイスの脳炎の牛の脳にBoAstV-CH15という未知のBoAstV株が同定された。 完全なゲノム系統発生比較は、boastv-CH13よりもovineアストロウイルス(OvAstV)とBoAstV-CH15の密接な関係を明らかにした(Seuberlich et al., 2016). Boastv−C H1 3およびBoastv−C H1 5との共感染もまた、1つの症例において文書化された(Seuberlich e t a l., 2016). 同じ年にドイツで、Schlottau et al. (2016)は、脳炎を有する牛において、OvAstVおよびBoAstV-CH15と最も密接に関連していた新規なアストロウイルス、すなわちBoAstV-BH89/14を報告した。 その後、BoAstV-CH13/Neurose1は、カナダ東部および西部のウシ脳炎の症例において2017年に同定された(Spinato et al. ら、2 0 1 7;Selimovic−Hamza e t a l.、2017b)。 2018年には、日本における非化膿性脳脊髄炎を伴うステアにおいて、北米および欧州のBoAstV-Neuros1/BoAstV-CH13と密接に関連する新規な神経侵襲性BoAstVが同定され、北米と欧州の株間の遺伝子型内組換えの発生が示唆された(Hirashima et al., 2018).
アストロウイルス関連脳炎の症例は北米、欧州、アジアで報告されているが、その存在は南半球では記録されていない。 ここでは、地理的分布と神経浸潤性アストロウイルスの遺伝的多様性を広げ、このウイルスがヨーロッパからアメリカに導入され、後にアジアに広
材料と方法
歴史とシグナル
2018年6月、ウルグアイのコロニアにある∼300ヘクタールの農場で37頭のグループの22ヶ月のホルスタインの雄牛が、異常な行動、目的のない歩行、旋回、運動失調、反復的で協調していない舌の動き、および臥床を含む進行性の神経学的歌を発症した。 群れは毎年恒例のオート麦の牧草地で放牧され、トウモロコシのサイレージが補充されました。 獣医開業医による脳リステリア症の推定臨床診断は、ペニシリンとストレプトマイシンによる治療を促したが、動物は3日間続く臨床経過後に自発的に死亡した。
病理学的検査、in situハイブリダイゼーション(ISH)および免疫組織化学(IHC)
雄牛の頭を枝肉から取り出し、診断作業のためにINIAの獣医診断研究所(Animal Health Platform)に提出した。 脳の半分、近位頚髄(C1)、三叉神経節および三叉神経の根、唾液腺、後咽頭リンパ節、中咽頭、食道、舌および骨格筋の短いセグメントは、10%中性緩衝ホルマリンに48-72グラムの汚れ。5Red assay kit(Cat#3 2 2 3 6 0,Advanced Cell Diagnostics,Hayward,C A,United States)およびBoastvプローブCatを使用して、superfrost Plus slids(Thermo Fisher Scientific,Pittsburgh,PA,United States)上のホルマリン固定、パラフィン埋め込み(FFPE)脳幹、大脳および小脳の5μ m切片に #406921. プローブは、ウイルスの領域5232-6180を標的とする20ZZペア(GenBank KF233994.1)で構成されている。 組織の各5μ mセクションは、プローブハイブリダイゼーションの前に熱およびプロテアーゼで前処理され、2時間40℃で製造業者の推奨に従って処理された。 信号の検証に使用される負のコントロールは、シリアルセクションと感染していない動物からの組織をプロービング上で実行される無関係な(GCコンテン スライドをヘマトキシリンで対比染色し、Ecomount(Biocare Medical,Concord,C A,United States)で装着した。
さらに、ウェストナイルウイルス(WNV、フラビウイルス)の同定のために、脳幹、大脳および小脳のFFPE切片においてIHCを行った(Palmieri et al. ら、2 0 1 1)、狂犬病ウイルス(リサウイルス)(Stein e t a l. ら、2 0 1 0)、およびChlamydia spp. (Giannitti e t a l.、2016年)。<7 7 5 5><7 1 3 3>Molecular Virology<9 4 1 2><1 5 9 9>Magmax Nucleic Acid Isolation Kit(登録商標)(Thermo Fisher Scientific)を用いて、凍結(−2 0℃)脳のプールされたサンプルから核酸抽出を達成した。 アストロウイルス検出のために、逆転写(R T)を、Revertaid Reverse Transcriptase(登録商標)(Thermo Fisher Scientific)およびrandom h examer primer(Qiagen)を用いて行った。 PCRは、mangomix(登録商標)(Bioline)およびアストロウイルスポリメラーゼ遺伝子の4 3 2ヌクレオチド断片を増幅するプライマー(Tse e t a l.,2 0 0 2)を使用してcDNAから実施した。, 2011). PCR産物を2%アガロースゲル中で可視化し、Purelink(登録商標)Quick Gel Extraction and PCR Purification Combo Kit(Invitrogen)を用いて精製し、Macrogen Incで配列決定した。 (ソウル、韓国)。 アストロウイルス全ゲノム増幅のためには、cDNAの観察のためのMaxima H Minus逆転写酵素(Thermo Fisher Scientific)およびoligo(D T)1 8、ならびにHirashimaらによって記載されるプライマーを用いたMangomix(登録商標)(Bioline)またはRanger DNA Polymerase(Bioline)が挙げられる。、2018年、使用された。 PCR産物を1-2%アガロースゲル中で可視化し、精製し、上記のように配列決定した。 配列の組み立ては、Seqman(Lasergene8,DNASTAR)を用いて行った。 牛、羊、豚、ヒトおよびミンクからの神経侵襲性アストロウイルスの二十から六の完全なゲノム配列、およびGenBankで利用可能な腸性ウシアストロウイルス, 2016). W−IQ−TREE1(Trifinopoulosら(1 9 9 9))。,2016)を用いて、配列進化の最良適合モデル(SYM+I+G4)を決定し、この場合に検出されたBoAstVのほぼ完全な配列とGenBankからダウンロードされた完全な配列との最尤系統樹を構築するために、bootstrapを用いてクレードのロバスト性を評価する統計的方法として使用した。 類似性プロットは、Simplotソフトウェア(Lole e t a l., 1999). ORF2のアミノ酸レベルでのp−距離は、MEGA7ソフトウェアを用いて推定した(Kumar e t a l., 2016).
さらに、BEAST v1.8.4パッケージを用いてベイズ系統地理学的解析を行った(Drummond et al.、2012年)を使用して、: BoAstV CH13/Neuros1系統の完全なコード領域、Orf1Ab(非構造遺伝子)、ORF2(構造遺伝子)、Orf1A(プロテアーゼ)および部分的なOrf1B(ポリメラーゼゲノム領域、カナダ株が利用可能であった)、GenBank(最後のアクセッションApril18、2019)で利用可能なすべての配列を用いて、進化速度、共通の祖先の年齢/年、および国(スイス、ウルグアイ、米国、カナダ、および日本)によるウイルス循環の最も可能性の高いルートを決定する。 データセット内の再結合の欠如は、再結合検出プログラム4を用いて決定された。 各アライメントに最もよく適合する置換モデルは、ベイズ情報基準(BIC)値を介してMEGA7ソフトウェアを使用して決定され、各データセットの時間構造はTempEst(Rambaut et al., 2016). Bayesスカイライン解析を用いた対数正規緩和分子時計を,使用した分子時計と合体樹プリオの異なる組み合わせの中からBayes因子によって選択した。 検出の国は形質として使用されました。 マルコフ連鎖モンテカルロの長さは100万世代であり、Tracer v1.6で評価された解析の収束を保証した。0、および事後確率を使用してクレードを評価しました。 最大クレード信頼性ツリー(MCCT)は、BEASTからTreeAnnotatorソフトウェアを使用して得られ、FigTree v1.4.3で視覚化されました。
最後に、前述のように、凍結脳から抽出されたDNAを、ウシヘルペスウイルス1および5(BHV-1および-5)の検出のためにPCRによって処理した(Ashbaugh et al., 1997).
細菌学
大脳および脳幹の新鮮なサンプルは、血液およびMacConkey寒天中の好気性細菌培養およびListeria monocytogenesの選択培養のために日常的に処理された(Al-Zoreky and Sandine、1990)。
結果と考察
本明細書に記載されている症例の臨床徴候および疫学的所見は、非特異的ではあるが、通常散発的であると記載されているウシアストロウィルス関連脳炎の他の症例に記載されているものと同様であった(Selimovic-Hamza et al. ら、2 0 1 6)、種々の神経学的欠損を伴う(Deiss e t a l. ら、2 0 1 7)、典型的には1日〜3週間の範囲の臨床徴候の持続時間を有する(Schlottau e t a l.,2 0 1 8)。 ら、2 0 1 6;Deiss e t a l. ら、2 0 1 7;Spinato e t a l. ら,2 0 1 7;Hirashima e t a l., 2018).
脳、脊髄のC1セグメント、および頭部の他の組織の肉眼的検査では、有意な肉眼的な解剖学的病変は明らかにされなかった。 組織学的には,終脳(大脳半球および海馬を含む),脳幹および脊髄の唯一の検査セグメントに影響を及ぼす中等度から重度のリンパ球性,組織球性および形質細胞性髄膜脳脊髄炎があった。 病変は主に灰白質および白質の限界領域に分布していた。 患部では血管周囲カフおよびリンパ球および組織球性炎症および隣接する神経障害にグリオーシスを伴う神経壊死/神経食が認められた。 影響を受けた壊死性ニューロンの衛星化があった(図1A–D)。 多巣性中等度小脳性レプトメニング炎が認められたが,小脳実質では病変の頻度ははるかに少なく,重症であった。 病巣内細菌はH&Eとグラム染色で発見されませんでした。 他の検査組織では有意な組織学的変化は認められなかった。

図1. 脳幹(A、B)および大脳皮質(C、D)における組織学的病変および大脳皮質(E、G)におけるBoAstV RNAの検出。 画像(A−D)は、H<9 0 9 4>Eで染色された脳の切片である。; 画像EとGは、ヘマトキシリンで対染色BoAstV特異的プローブを用いた発色ISHを用いたハイブリダイゼーションを示す大脳皮質のセクションであり、画像Fは、ヘマトキシリンで対染色された大脳皮質DapBプローブ(陰性対照)のシリアルセクションである。 (A)周囲腔は、隣接する神経細胞にも浸潤する炎症性細胞(主にリンパ球および組織球)によって著しく拡張される。 (B)中心のニューロンにhypereosinophilic perikaryonおよびkaryorrhexis(壊死)があり、ニューロンボディはグリア(satellitosis)および炎症性細胞の増加された数によって囲まれています。 (C)レプトメニンゲは、リンパ球および組織球によって浸潤される。 (D)多数のhypereosinophilic(necrotic)ニューロンおよびperivascularリンパ球の袖口が付いている大きい容器が付いている大脳皮質の領域。 (E)大脳皮質のシリアルセクションでは、豊富な細胞質内BoAstV RNAラベリングは、負の対照プローブ(F)を使用してハイブリダイゼーションでは存在しないソーマと神経延長(E、G)の神経細胞質内の強い、顆粒状の赤色色原体沈着によって描かれている。
中枢神経系の組織学的検査で神経浸潤性ウイルス感染が疑われた。 多くの反芻動物の神経病原体が人獣共通感染症であるため、脳炎を有する牛が懸念されている(Cantile and Youssef、2016); したがって、脳炎の診断は、可能な場合には感染性病原体をスクリーニングするために広範な実験室試験を促すべきである。 本明細書に記載の場合、WNV、狂犬病ウイルスおよびクラミジア属に対するIHCs。、およびBHV-1および-5のPCRはすべて陰性であり、病原菌は脳組織から培養されなかった。 雄牛は<2歳であり、脳幹に海綿状の変化は観察されなかったため、ウルグアイでは報告されていない成牛のエキゾチックな病気であるウシ海綿状脳症(BSE)の試験は行われなかった。 さらに、BSEは炎症性ではない(Cantile and Youssef、2016)。
BoAstV-Neurose1から生成されたプローブを用いてIn situハイブリダイゼーションを行い、大脳半球および海馬のニューロンの細胞質内に豊富で限定されたプローブハイブリダイゼーションがあった(図1E–G)。 これらの領域では、プローブハイブリダイゼーションは、グリア細胞、または血管周囲袖口の炎症細胞で検出可能なないプローブハイブリダイゼーションと、壊死性ニューロンとグリア症の領域と共局在した。 Ishによるウイルス核酸は小脳には検出されず,実質には炎症性病変は最小限であったが中等度のレプトメニング炎,または重度の炎症を伴う切片を含む脳幹であった。 これは、地形的に、ISHによるウイルス分布の検出が、ウシにおけるBoastv−C H1 3/Neuros1関連脳炎の症例で時折記載されている、検査されたセクションにおける脳炎よ,2017a,b). 脳の病変領域におけるウイルスRNA検出のこの時折の欠如の理由は、以前に示唆されたように、ISHの検出限界、または死亡時までの脳の炎症領域におけるウ、2017b)。 予想されるように、陰性対照として使用された脳組織において、ishによってプローブハイブリダイゼーションは検出されなかった。
アストロウイルスはRT-PCRにより脳内に検出された。 ほぼ完全なゲノム配列解析は、CH13/Neuros1クレード内のママストロウイルス株を明らかにし、我々はBoAstV-Neuro-Uyと命名し、配列はaccession番号MK386569の下でGenBankに寄託された。 系統解析では、最も知られている神経侵襲性アストロウイルスを含むバージニア/ヒトミンクオビン(VA/HMO)クレード内の他の神経侵襲性アストロウイルスとの近 ら,2 0 1 8;Reuterら,2 0 1 9., 2018). BoAstV-Neuro-Uyのほぼ完全な配列は、長さが6427bpであり、Kagoshimasr28-462株と94%の配列同一性を有する。 BoAstV-Neuro-Uyは系統CH13/Neuros1の他の株と同様の特徴を持っています:51ntの5’UTR領域、861アミノ酸(aa)のOrf1A(プロテアーゼ)、523aa(RNA依存性RNAポリメラーゼ)のOrf1B、および758aa(キャプシドタンパク質)のORF2。 残念なことに、3’UTRは配列決定できなかったが、オリゴ(d T)1 8をcDNAを得るために使用したので、ポリ(A)尾部が存在すると推定される。 さらに、リボソームフレームシフトシグナルであるヘプタマー AAAAAAC配列が存在する。 ORF2のアミノ酸レベルでのP距離は、CH13/Neuros1クレードへのこの株の割り当てを確認した。 P-距離<0.Mamastrovirus−1 3は、他の著者によって最近提案されている(Donato and Vijaykrishna,2 0 1 7;Hirashima e t a l.,2 0 1 7)。 2018年現在、ICTVによる明確な種の割り当ては保留されている。 BoAstV-Neuro-Uyから生成されたISHに使用されるプローブは、BoAstV-Neuro-Uyと92.7%の配列同一性を有していた。
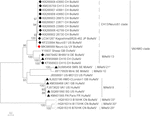
図2. 最尤法による全長ヌクレオチド配列の系統発生解析。 BoAstV-Neuro-Uyは赤い菱形でマークされています。 牛のようなneuroinvasiveアストロウイルスの他の順序は黒い菱形と印が付いています、非牛のようなneuroinvasiveアストロウイルスからの順序は黒い三角形と印が付いています、そして腸性牛のようなアストロウイルスからの順序は白い円と印が付いています。 ママストロウイルス種が示されており、アスタリスクはICTVによってまだ認識されていない種を示しています。 ICTVによってまだ種が割り当てられていないC H1 3/Neuros1クレード内の株は、破線で同定される。 ブートストラップ値はノードで示されます。 CH、スイス、JP、日本、米国、アメリカ合衆国; Uy、ウルグアイ;GB、イギリスのイギリス;DE、ドイツ;SE、スウェーデン;FR、フランス;CN、中国。 ス;HUASTV、ヒトアストロウイルス;Miastv、ミンクアストロウイルス;HUASTV、ヒトアストロウイルス;Huastv、ヒトアストロウイルス;Huastv、ヒトアストロウイルス;Huastv、ヒトアストロウイルス;Huastv、ヒトアストロウイルス;Huastv、ヒトアストロウイルス;Huastv
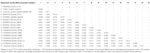
表1. ウシとオビのママストロウイルス-13の配列とCH13/Neuros1クレード内の株の間の完全なORF2領域のアミノ酸レベルでの進化の発散の推定は、まだウイルスの分類学(ICTV)に関する国際委員会によって種に割り当てられていない。
神経病理学的検査およびアストロウイルス核酸およびタンパク質の検出に基づく研究は、アストロウイルス感染と牛の神経疾患および病変との間に因果関係があると結論づけている(Selimovic-Hamza et al. ら、2 0 1 7a;Reuterら、2 0 1 8a., 2018). 我々の知る限りでは、アストロウイルス関連脳炎はまだ実験的に再現されていない。 これは臨床例からの神経浸潤性アストロウイルスの分離を必要とするが,これは我々の症例では試みられなかった。
この場合のBoAstV-Neuro-Uyの発生源は特定できませんでした。 しかし、牛は広範な屋外条件下で飼育されているため、貯水池の牛や野生動物を考慮する必要があります。 影響を受けた動物は、2018年2月に他の9頭とともに購入され、農場に移されました。 残念なことに、所有者は、プロパティ内の他の動物のさらなるサンプリングとテスト、およびより詳細な疫学的調査を拒否しました。 グループ内の他の動物のいずれも、獣医開業医が最後に接触した8月の2018の時点で神経学的疾患を発症していなかった。 スイスにおけるアストロウイルス関連脳炎の症例については、冬の初めから春の終わりまでの季節性が示唆されている(Selimovic-Hamza et al., 2016). 興味深いことに、ここに記載されているケースは、南半球の秋-冬の移行期に対応して、月に発生しました。
北米では神経栄養性アストロウイルスが同定されている(Li et al. ら,2 0 1 3;Spinato e t a l. ら、2 0 1 7)、欧州(Bouzalas e t a l. ら、2 0 1 4)およびアジア(Hirashima e t a l. 2018年)、彼らの存在は南半球では報告されていないので、この通信はアストロウイルス関連脳炎の地理的分布を広げる。 ウルグアイで検出されたウイルス株は、ヨーロッパ、北米、またはアジアに由来している可能性があるかどうかを評価するために、我々は進化速度を推定し、GenBankで利用可能な神経侵襲的BoAstVシーケンスを使用して系統地理学的解析を行った。 完全な符号化領域を用いて推定された進化速度は4であった。27×10-4(95%最高確率密度-HPD–,2.19-6.46×10-4)ヌクレオチド置換/部位/年であり、これはRNAウイルスについて予想される(Jenkins et al. ら、2 0 0 2)、しかし、腸内ヒトアストロウイルスについて推定されたものよりも低い(Babkin e t a l., 2012, 2014). Orf1Ab領域は、完全なコード領域と同様の進化速度(4.20×10–4、95%HPD1.66-6.46×10-4置換/サイト/年)を示し、一方、Orf1A(2.92×10-4、95%HPD1.19×10-6-6.46×10-4置換/10-6-579×10-4置換/サイト/年)はわずかに速い進化速度を示し、部分ポリメラーゼゲノム領域(Orf1B)はわずかに遅い進化速度(5.39×10-4、95%HPD6.41×10-7-1.10×10-3置換/サイト/年)を示した。
完全なコード領域を持つ系統解析によって決定され、MCTTに示されているように(図3)、共通の祖先を持つ参照株に基づいて二つのサブ系統(CH13とNeurose1)があ これらの亜系統(系統CH13/Neurose1)の最も最近の共通の祖先は、およそ1885年(95%HPD、1794年-1940年)にヨーロッパで発生しました。 1900年代の初めに、二つのサブ系統が分岐し、CH13サブ系統はヨーロッパで循環し続け、Neurose1サブ系統はアメリカとアジアに広がった。 最も可能性の高いシナリオは、1921年頃(95%HPD、1849-1967)にヨーロッパからウルグアイに導入されたNeurose1サブ系統が、おそらく家畜貿易を通じて北米に、後に日本に広 GenBankで利用可能な配列数の制限のために、分析に偏っている可能性があり、完全なコード領域を使用して得られた結果は、より多くの株(すなわち、カナダ株) すべての分析において、最も可能性の高いシナリオは、ウルグアイへのウイルスの導入がヨーロッパから発生したことである(補足図S1A–D)。 さらに、Orf1Abおよび部分ポリメラーゼゲノム領域(Orf1B)で得られたこの導入の推定日(補足図S1A、D)は、完全なコード領域で得られたものと同様であったが、ORF2およびOrf1Aで得られた導入の推定日は以前であったが、より広い95%HPD間隔(補足図S1B、C)であった。 副系統Neurose1をヨーロッパからカナダに直接導入し、その後米国および日本に普及させたことも妥当であり、補足図S1Dに示されている。

図3. 全長符号化領域の解析により得られる最大クレード信頼性ツリー。 枝の色は祖先が循環した最も可能性の高い国を表し、枝に事後確率値が表示され、各ノードの数字は95%のHPD間隔で各クレードの起源の年を表します。 副系統はラベルで示される。
地理的分布、病原性メカニズム(特に伝播と侵入のメカニズム)、分子疫学、および神経侵襲性アストロウイルスの種間伝達の可能性を評価するためには、さらなる調査が必要である。
データの可用性
この研究のために生成されたデータセットは、GenBank、MK386569にあります。
著者の貢献
FG、RDC、MCはこの研究の概念に貢献しました。 FgとRDCは病理学的検査とサンプリングを行った。 PPはinsituハイブリダイゼーションを行った。 FUは免疫組織化学を行った。 LM,RC,MCは分子ウイルス学的試験を行った。 MCは配列および系統地理学的解析および関連する図を行った。 FgとPPは組織像を得た。 MFは細菌培養を行った。 FGとMCは原稿の最初の草案を書いた。 RDC、PP、FU、LM、MF、RCは原稿のセクションを書いた。 すべての著者は原稿の改訂に貢献し、提出されたバージョンを読んで承認しました。
資金
この作品は、INIAからの助成金PL-015N-15156と、「Comisión Sectorial de Investigación Científica」(CSIC)からの「Programa de Iniciación a la Investigación2017」からの158によって資金提供されました。 MCとRDCは、博士号奨学金を通じて、それぞれ「国立研究・技術革新機関」(ANII)とINIAからの支援を認めています。 FGは、モビリティグラントMOV_CA_2018_1_150021を通じてANIIからのサポートを認めています。
利益相反声明
著者らは、この研究は潜在的な利益相反と解釈される可能性のある商業的または財政的関係がない場合に行われたと宣言している。
レビュアー TSは、著者PPの一人との過去の共著者をハンドリングエディタに宣言しました。
謝辞
著者は、INIAのYisell PerdomoとCecilia Monesiglio、CAHFSのKaren SverlowとJuliann Beingesserに技術支援を感謝しています。
補足資料
この記事の補足資料はオンラインで見つけることができます: https://www.frontiersin.org/articles/10.3389/fmicb.2019.01240/full#supplementary-material
図S1/全長Orf1A B(A)、全長ORF2(B)、全長Orf1A(C)、および部分Orf1B(D)の分析によって得られた最大クレード信頼性木(Mcct)。 枝の色は祖先が循環した最も可能性の高い国を表し、枝には事後確率値が表示され、各ノードの数字は95%HPD間隔の各クレードの起源年を表します。 副系統はラベルで示される。
- ^http://iqtree.cibiv.univie.ac.at
およびSandine,W. 食糧からのListeria monocytogenesの分離のための非常に選択的な媒体。 Appl. エンビロン ミクロビオール 56, 3154–3157.
ることができると考えられている(1 9 9 7年)。 入れ子にされたポリメラーゼの連鎖反応を使用して牛のようなヘルペスウイルス1および5の取除くことそして潜伏の特定の検出。 J.Vet. Diagn。 投資する。 9, 387–394. 土井:10.1177/104063879700900408
PubMed要約/CrossRef全文/Google Scholar
Babkin,I.V.,Tikunov,A.Y.,Sedelnikova,D.A.,Zhirakovskaia,E.V.,And Tikunova,N.V.(2014). Hastv-2およびHastV-4完全ゲノムに基づく組換え解析。 感染した ジュネット エヴォル 22, 94–102. ドイ:10.1016/j.meegid.2014.01.010
PubMed要約/CrossRef全文/Google Scholar
Babkin,I.V.,Tikunov,A.Y.,Zhirakovskaia,E.V.,Netesov,S.V.,And Tikunova,N.V.(2012). ヒトアストロウイルスの高い進化率。 感染した ジュネット エヴォル 12, 435–442. ドイ:10.1016/j.meegid.2012.01.019
PubMed要約/CrossRef全文/Google Scholar
とができるようになりました。 ウイルスメタゲノミクスを用いた振とうミンク症候群に罹患しているミンクの脳組織における新規アストロウイルスの検出。 J.Clin. ミクロビオール 48, 4392–4396. 土井:10.1128/JCM.01040-10
PubMed要約/CrossRef全文/Google Scholar
ボロス、アルベルト、M.、パンコヴィックス、P.、Bíró、H.、Pesavento、P.A.、Phan、T.G.、ら。 (2017). ハンガリー、離乳豚の間で脳脊髄炎、脱力感、および麻痺に関連する神経浸潤性アストロウイルスの発生。 エメログ 感染した Dis。 23, 1982–1993. doi:10.3201/eid2312.170804
PubMed要約/CrossRef全文/Google Scholar
Boujon,C.L.,Koch,M.C.,Wüthrich,D.,Werder,S.,Jakupovic,D.,Bruggmann,R.,et al. (2017). ヒツジおよび牛の脳炎と関連付けられるアストロウイルスの交差種伝達の徴候。 エメログ 感染した Dis。 23, 1604–1608. ドイ:10.3201/eid2309.170168
PubMed Abstract/CrossRef Full Text/Google Scholar
Bouzalas,I.G.,Wüthrich,D.,Selimovic-Hamza,S.,Drögemüller,C.,Bruggmann,R.,And Seuberlich,T.(2016). 脳炎関連ウシアストロウイルスのフルゲノムベースの分子特性評価。 感染した ジュネット エヴォル 44, 162-168. ドイ:10.1016/J.meegid。06/052/2016
PubMed要約/CrossRef全文/Google Scholar
Bouzalas,I.G.,Wüthrich,D.,Walland,J.,Drögemüller,C.,Zurbriggen,A.,Vandevelde,M.,et al. (2014). ヨーロッパのnonsuppurative脳炎の牛のNeurotropicアストロウイルス。 J.Clin. ミクロビオール 52, 3318–3324. 土井:10.1128/JCM.01195-14
PubMed要約/CrossRef全文/Google Scholar
ることができます。 Jubb,Kennedy,And Palmer’s Pathology of Domestic Animalsの「神経系」、第6版、編。 M.G.Maxie(セントルイス,MO:Elsevier),205-406.
とができるようになりました。 アストロウイルス関連脳炎を有する牛の神経学的臨床徴候。 J.Vet. インターン メッド 31, 1209–1214. 土井:10.1111/jvim.14728
PubMed要約/CrossRef全文/Google Scholar
ることができます。 哺乳類および鳥のアストロウイルスの広いホストの範囲そして遺伝の多様性。 9:E102. doi:10.3390/v9050102
PubMed要約/CrossRef全文/Google Scholar
とができるようになりました。 Bayesian phylogenetics with BEAUti and THE BEAST1.7. モル バイオル エヴォル 29, 1969–1973. doi:10.1093/molbev/mss075
PubMed要約/CrossRef全文/Google Scholar
Giannitti,F.,Anderson,M.,Miller,M.,Rowe,J.,Sverlow,K.,Vasquez,M.,et al. (2016). クラミジアのpecorum:散発的なカプリンの中絶の胎児および胎盤がある損害。 J.Vet. Diagn。 投資する。 28, 184–189. 土井: 10.1177/1040638715625729
PubMed Abstract|CrossRef全文|Google Scholar
平嶋陽菜、岡田大、柴田大、吉田大、藤園大、大松大、他… (2018). 日本における非化膿性脳脊髄炎を伴うニホンブラックステアで検出された新規神経栄養性ウシアストロウイルスの全ゲノム解析。 アーチ ヴィロール 163, 2805–2810. ドイ:10.1007/s00705-018-3898-3
PubMed Abstract|CrossRef全文|Google Scholar
ウイルスの分類に関する国際委員会(2018)。 マスター種リスト(MSL32)、バージョン1。 利用可能な場所:https://talk.ictvonline.org/files/master-species-lists/m/msl/7185(2019年3月アクセス)。
Jenkins,G.M.,Rambaut,A.,Pybus,O.G.,And Holmes,E.C.(2002). RNAウイルスにおける分子進化の速度: 定量的な系統発生分析。 J.モル。 エヴォル 54, 156–165. ドイ:10.1007/s00239-001-0064-3
PubMed Abstract|CrossRef全文|Google Scholar
とができると考えています。 MEGA7:より大きなデータセットのための分子進化遺伝学解析バージョン7.0。 モル バイオル エヴォル 33, 1870–1874. doi:10.1093/molbev/msw054
PubMed要約/CrossRef全文/Google Scholar
Li,L.,Diab,S.,McGraw,S.,Barr,B.,Traslavina,R.,Higgins,R.,et al. (2013). 牛の神経学的疾患に関連する発散性アストロウイルス。 エメログ 感染した Dis。 19, 1385–1392. doi:10.3201/eid1909.130682
PubMed要約/CrossRef全文/Google Scholar
Lole,K.S.,Bollinger,R.C.,Paranjape,R.S.,Gadkari,D.,Kulkarni,S.S.,Novak,N.G.,et al. (1999). サブタイプ間組換えの証拠とインドのサブタイプC感染seroconvertersから全長ヒト免疫不全ウイルス1型ゲノム。 J.ヴィロール 73, 152–160.
Naccache,S.N. ら、Peggs,K.S.,Mattes,F.M.,Phadke,R.,Garson,J.A.,Grant,P.,et al. (2015). 公平な次世代の配列決定による脳炎のimmunocompromised大人のneuroinvasiveアストロウイルスの伝染の診断。 クリン 感染した Dis。 60, 919–923. doi:10.1093/cid/ciu912
PubMed要約/CrossRef全文/Google Scholar
Palmieri,C.,Franca,M.,Uzal,F.,Anderson,M.,Barr,B.,Woods,l.,et al. (2011). Psittaciformesにおける西ナイルウイルス感染の病理学的所見と免疫組織化学的所見。 獣医さん パトル 48, 975–984. 土井:10.1177/0300985810391112
PubMed Abstract/CrossRef Full Text|Google Scholar
Pfaff,F.,Schlottau,K.,Scholes,S.,Courtenay,A.,Hoffmann,B.,Höper,D.,et al. (2017). 国内のヒツジにおける脳炎および神経節炎に関連する新規アストロウイルス。 トランスバウンド エメログ Dis。 64, 677–682. 土井:10.1111/tbed.12623
PubMed要約/CrossRef全文/Google Scholar
Quan,P.L.,Wagner,T.A.,Briese,T.,Torgerson,T.R.,Hornig,M.,Tashmukhamedova,a.,et al. (2010). Xリンクagammaglobulinemiaの男の子のアストロウイルス脳炎。 エメログ 感染した Dis。 16, 918–925. doi:10.3201/eid1606.091536
PubMed要約/CrossRef全文/Google Scholar
とができるようになりました。 テンペスト(formely Path-O-Gen)を用いてヘテロクロナス配列の時間構造を探索する。 ウイルスエヴォル… 2:vew007. doi:10.1093/ve/vew007
PubMed要約/CrossRef全文/Google Scholar
ることができます。 人間および動物のneurovirulentアストロウイルスの伝染と関連付けられるNonsuppurative(無菌)髄膜脳脊髄炎。 クリン ミクロビオール Rev.31,e00040-18. ドイ:10.1128/CMR.00040-18
PubMed Abstract/CrossRef Full Text|Google Scholar
Schlottau,K.,Schulze,C.,Bilk,S.,Hanke,D.,Höper,D.,Beer,M.,et al. (2016). 脳炎の牛の新しい牛のようなアストロウイルスの検出。 トランスバウンド エメログ Dis。 63, 253–259. 土井:10.1111/tbed.12493
PubMed要約/CrossRef全文/Google Scholar
セリモビッチ-ハムザ,S.,ブジョン,C.L.,ヒルベ,M.,Oevermann,A.,And Seuberlich,T.(2017a). 神経疾患牛におけるウシアストロウイルスCH13/Neuros1感染の頻度と病理学的表現型:因果関係の評価に向けて。 9月12日にメジャー契約を結んでアクティブ-ロースター入り。 doi:10.3390/v9010012
PubMed要約/CrossRef全文/Google Scholar
セリモビッチ-ハムザ,S.,サンチェス,S.,フィリベルト,H.,クラーク,E.G.,とSeuberlich,T.(2017b). カナダ西部の神経学的疾患を有する肥育牛におけるウシアストロウイルス感染。 できます。 獣医さん J.58,601-603.
セリモビッチ-ハムザ,S.,Bouzalas,I.G.,Vandevelde,M.,Oevermann,A.,And Seuberlich,T.(2016). ヨーロッパの散発性ウシ脳炎の歴史的症例におけるアストロウイルスの検出、スイス1958-1976。 フロント 獣医さん サイ… 3:91. ドイ:10.3389/fvets.2016.00091
PubMed要約/CrossRef全文/Google Scholar
ゼーベリッヒ,T.,ヴュートリッヒ,D.,セリモビッチ-ハムザ,S.,ドレーゲミュラー,C. ら,oevermann,A.,Bruggmann,R.,et a l. (2016). 牛における第二脳炎関連アストロウイルスの同定。 エメログ 微生物が感染する。 5:e5. 土井:10.1038/2016.5
PubMed要約/CrossRef全文|Google Scholar
とができるようになりました。 カナダ東部の牛のような非化膿性脳炎の場合の牛のようなアストロウイルスの同一証明。 できます。 獣医さん J.58,607-609.
とができるようになりました。 国内および野生生物種の中枢神経系内の狂犬病ウイルスの免疫組織化学的研究。 獣医さん パトル 47, 630–633. 土井: 10.1177/0300985810370013
PubMed Abstract|CrossRef全文|Google Scholar
とができるようになりました。 W-IQ-TREE:最尤分析のための高速オンライン系統発生ツール。 核酸Res.44,W232-W235. doi:10.1093/nar/gkw256
PubMed要約/CrossRef全文/Google Scholar
Tse,H.,Chan,W.M.,Tsoi,H.W.,Fan,R.Y.,Lau,C.C.,Lau,S.K.,et al. (2011). 牛のようなアストロウイルスの再発見そしてゲノムの性格描写。 J-ジェン-ヴィロル 92(Pt8),1888-1998. ドイ:10.1099/vir。0.030817-0
PubMed Abstract/CrossRef Full Text|Google Scholar