生物学専攻II
学習成果
- ウイルスの過去および新たな分類システムを理解する
ほとんどのウイルスはおそらく異なる祖先から進化したため、科学者が原核細胞と真核細胞を分類するために使用した体系的な方法はあまり有用 ウイルスが異なる生物の”残党”を表す場合、ゲノム分析やタンパク質分析でさえ有用ではありません。 どうして? ウイルスは、彼らがすべて共有する共通のゲノム配列を持っていないので。 例えば、原核生物の系統発生を構築するのに非常に有用な16S rRNA配列は、リボソームのない生き物には役に立たない! 生物学者は過去にいくつかの分類システムを使用してきました。 ウイルスは、最初は共有形態によってグループ化されました。 その後、ウイルスの群は、それらが含有する核酸の種類、DNAまたはRNA、およびそれらの核酸が一本鎖または二本鎖であるかどうかによって分類された。 彼らはウイルスの文字の異なるセットに基づいていたので、しかし、これらの以前の分類方法は、異なるウイルスをグループ化しました。 今日最も一般的に使用されている分類方法は、ボルチモア分類スキームと呼ばれ、メッセンジャー RNA(mRNA)がウイルスの各特定のタイプで生成される方法に基
過去の分類システム
ウイルスには、ウイルスゲノム、カプシドの種類、エンベロープウイルスのエンベロープ構造など、分類できる要素はわずかです。 これらの要素のすべては、過去にウイルス分類のために使用されてきた(表1および図1)。 ウイルスゲノムは、遺伝物質の種類(DNAまたはRNA)およびその組織(一本鎖または二本鎖、線状または環状、およびセグメント化または非セグメント化)が異 いくつかのウイルスでは、複製に必要な追加のタンパク質は、ゲノムに直接関連付けられているか、ウイルスカプシド内に含まれています。
| 表1. ゲノム構造とコアによるウイルス分類 | |
|---|---|
| コア分類 | 例 |
| RNA | 狂犬病ウイルス、レトロウイルス |
| DNA | ヘルペスウイルス、天然痘ウイルス |
| 一本鎖 | 狂犬病ウイルス、レトロウイルス |
| 二本鎖 | ヘルペスウイルス、天然痘ウイルス |
| リニア | 狂犬病ウイルス、レトロウイルス、ヘルペスウイルス、天然痘ウイルス |
| サーキュラー | パピローマウイルス、多くのバクテリオファージ |
| セグメント化されていない:ゲノムは、遺伝物質の単一のセグメントで構成されています | パラインフルエンザウイルス |
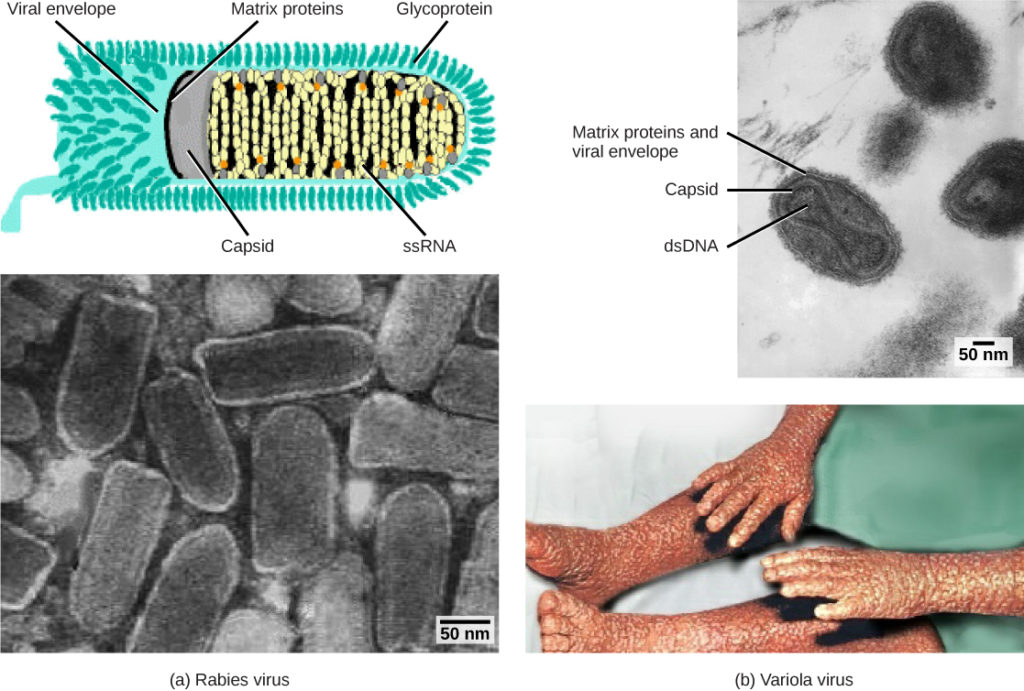
図1. ウイルスは、そのコア遺伝物質とカプシド設計に基づいて分類されます。 (a)狂犬病ウイルスは一本鎖RNA(ssRNA)コアとエンベロープされたヘリカルカプシドを有し、(b)天然痘の原因物質であるバリオラウイルスは二本鎖DNA(dsDNA)コアと複合カプシドを有する。 (クレジット”狂犬病図”:CDCによる作業の変更;”狂犬病マイクログラフ”:博士フレッド*マーフィー、CDCによる作業の変更;クレジット”小痘マイクログラフ”:博士フレッド*マーフィー、シルビア*ウィットフィールド、CDCによる作業の変更;クレジット”天然痘写真”:CDCによる作業の変更; Matt Russellのスケールバーデータ)
ウイルスはまた、それらのカプシドの設計によって分類することができる(表2および図2)。 カプシドは裸の二十面体、包まれた二十面体、包まれた螺旋形、裸の螺旋形、および複合体として分類される。 遺伝物質のタイプ(DNAまたはRNA)およびその構造(一本鎖または二本鎖、線状または環状、およびセグメント化または非セグメント化)を使用して、ウイル
| 表2. カプシド構造によるウイルス分類 | |
|---|---|
| カプシド分類 | 例 |
| 裸の二十面体 | 型肝炎ウイルス、ポリオウイルス |
| 包接二十面体 | エプスタインバーウイルス、単純ヘルペスウイルス、風疹ウイルス、黄熱病ウイルス、HIV-1 |
| インフルエンザウイルス、おたふく風邪ウイルス、麻疹ウイルス、狂犬病ウイルス | |
| ネイキッドヘリカル | タバコモザイクウイルス |
| 多くのタンパク質との複合体; いくつかは、二十面体構造とヘリカルカプシド構造の組み合わせを持っています | ヘルペスウイルス、天然痘ウイルス、B型肝炎ウイルス、T4バクテリオファージ |

図2。 種々のウイルスの透過型電子顕微鏡写真はその構造を示す。 (A)ポリオウイルスのカプシドは裸の二十面体である;(b)エプスタイン-バーウイルスのカプシドは包まれた二十面体である;(c)流行性耳下腺炎のウイル (クレジットa:Dr.Fred Murphy,Sylvia Whitfieldによる作業の変更;クレジットb:Liza Grossによる作業の変更;クレジットc:Dr.F.A.Murphy,CDCによる作業の変更;クレジットd:USDA ARSによる作業の: ケープタウン大学、南アフリカ、NASAの医学微生物学部門であるLinda Stannardによる作業の修正。)
ボルチモア分類
最も一般的に使用されているウイルス分類システムは、1970年代初頭にノーベル賞を受賞した生物学者デイビッド-ボルチモアによって開発されたものであり、上記の形態学や遺伝学の違いに加えて、ボルチモア分類スキームは、ウイルスの複製サイクル中にmRNAがどのように産生されるかに応じてウイルスを分類する。
グループIウイルスは、ゲノムとして二本鎖DNA(dsDNA)を含んでいます。 それらのmRNAは、細胞DNAとほぼ同じ方法で転写によって産生される。
グループIIウイルスは、ゲノムとして一本鎖DNA(ssDNA)を持っています。 それらは、mRNAへの転写が起こる前に、それらの一本鎖ゲノムをdsDNA中間体に変換する。
グループIIIウイルスはdsrnaをゲノムとして使用します。 鎖は分離し、そのうちの一つは、ウイルスによってコードされるRNA依存性RNAポリメラーゼを使用してmRNAの生成のための鋳型として使用される。
グループIVウイルスは、正の極性を持つゲノムとしてssRNAを持っています。 正の極性は、ゲノムRNAがmRNAとして直接作用することができることを意味する。 複製中間体と呼ばれるdsRNAの中間体は、ゲノムRNAをコピーする過程で作られます。 これらの中間体から負極性(正鎖ゲノムRNAに相補的)の複数の完全長RNA鎖が形成され、次いで、完全長ゲノムRNAおよびより短いウイルスmRNAの両方を含む正極性を有するRNAの産生のための鋳型として働くことができる。
グループVウイルスは負の極性を持つssRNAゲノムを含み、その配列はmRNAに相補的であることを意味する。 グループIVウイルスと同様に、dsrna中間体は、ゲノムのコピーを作成し、mRNAを生成するために使用されます。 この場合、負鎖ゲノムはmRNAに直接変換することができる。 さらに、完全長の正のRNA鎖は、負の鎖ゲノムの産生のための鋳型として機能するように作られる。
グループVIウイルスは、酵素逆転写酵素を用いてdsDNAに変換する必要がある二倍体(二つのコピー)ssRNAゲノムを持っています; 次いで、dsDNAは宿主細胞の核に輸送され、宿主ゲノムに挿入される。 そして、宿主ゲノムに組み込まれたウイルスDNAの転写によってmRNAを産生することができる。
グループVIIウイルスは、部分的なdsDNAゲノムを有し、mRNAとして作用するssRNA中間体を作るが、ゲノム複製に必要な逆転写酵素によってdsdnaゲノムに変換される。 ボルチモア分類における各グループの特徴は、各グループの例とともに表3に要約されている。
| 表3. ボルチモア分類 | |||
|---|---|---|---|
| グループ | 特徴 | mRNA産生のモード | 例 |
| I | 二本鎖DNA | mRNAはDNAテンプレートから直接転写されます | 単純ヘルペスウイルス(ヘルペスウイルス) |
| II | 一本鎖DNA | RNAが転写される前にDNAが二本鎖形態に変換される | イヌパルボウイルス(パルボウイルス) |
| III | 二本鎖RNA | mRNAはから転写される RNAゲノム | 小児胃腸炎(ロタウイルス) |
| IV | 一本鎖RNA(+) | ゲノムはmRNAとして機能する | 風邪(ピルコルナウイルス) |
| V | 一本鎖RNA(−) | rnaゲノムからmRNAが転写される | 狂犬病(ラブドウイルス) |
| VI | 逆転写酵素を有する一本鎖RNAウイルス | 逆転写酵素はRNAゲノムからDNAを作り、DNAは宿主ゲノムに組み込まれる; mRNAは、組み込まれたDNA | ヒト免疫不全ウイルス(HIV)から転写されます) |
| VII | 逆転写酵素を有する二本鎖DNAウイルス | ウイルスゲノムは二本鎖DNAであるが、ウイルスDNAはRNA中間体を介して複製され、RNAはmRNAとして直接またはmRNAを作) | |