Cell Biology06:The Cytoskeleton Part II:Tubulin
これらはHarvard ExtensionのCell Biologyコースの第6講義のノートです。
先週はアクチンで作られたマイクロフィラメントをカバーしました。 今週:チューブリンで作られた微小管。 なぜ、あなたは尋ねる、細胞骨格は二つの別々のシステムを必要としていますか? マイクロフィラメントは個々のサブユニットの単なる鎖であるのに対し、微小管は文字通り(すぐに見るように)二量体で作られた鎖で作られた壁を持つ中空の円筒であると考えてください。 これらの異なる構造は、異なる機能を意味します: マイクロフィラメントは、より簡単に自発的に形成することができ、異なるネットワーク形状を形成し、細胞の異なる部分を接続するために(Arp2/3およ 微小管は、形成するために核形成に大きく依存しており、基本的には中心体を細胞の周囲に接続するスポークネットワークである。 実際には、マイクロフィラメントは細胞皮質に豊富であり、収縮運動および細胞運動に大きく関与しているが、微小管はオルガネラの組織化に最も関与しており、前行性および逆行性輸送のトラックとして機能している。
微小管
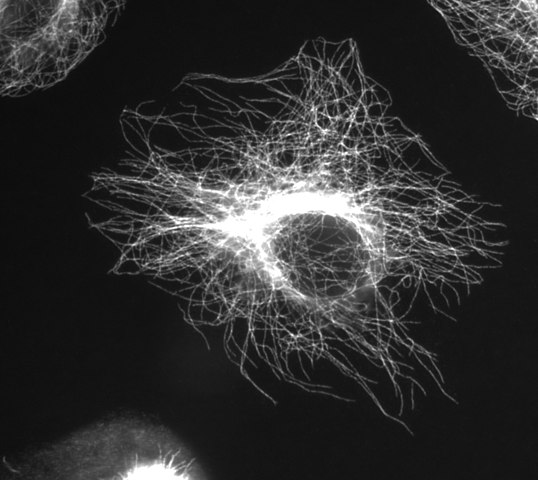
(ウィキメディア-コモンズのユーザー Jeffrey81のおかげで画像)
ウィキペディアに加えて、NCBIで利用可能なLodishとCooper細胞生物学の教科書の以前の版も優れた参考文献
微小管の管状構造は、マイクロフィラメントよりも頑丈であり、多くの重い義務の引っ張りと押しを可能にする。 頑丈ですが、微小管は温度に依存します-4℃に冷却すると解重合し、GTPが利用可能であれば37℃に加熱すると再び再重合します。
微小管の基本的な構成要素は、α-tubuilin/β-tubulin二量体(それぞれTUBA_遺伝子およびTUBB_遺伝子によってコードされる)である。 個々のチューブリンタンパク質はそれぞれ-55kdaの重量を量ります、従って-110da/アミノ酸で500アミノ酸の順序にあります。 Α-およびβ-チューブリンは両方ともGTPに結合するが、αはほとんどそれを永遠に保持するだけであるのに対し、βではGDPに交換または加水分解され、再び新しいGTPと交換することができる。
連続したα/β二量体の鎖がプロトフィラメントを構成し、13個のプロトフィラメントが円柱の中に並んで配置されて微小管を構成する。 プロトフィラメントの間のスペースは”継ぎ目”と呼ばれます。 微小管全体は直径約25nmである。 微小管についての細胞セグメントの内部生活の中で、その構造(二量体とプロトフィラメントで作られた)を見ることができます:
各二量体の中で、βサブユニットは(+)末端であり、重合に適しており、αは(-)末端であり、解重合に適している。 微小管は、核形成、伸長および定常状態であるマイクロフィラメントと基本的に同じ三つのステップで形成される。 しかし、マイクロフィラメントとは異なり、それらはそれ自身で容易に核形成しないので、核形成には微小管組織化センター(Mtoc)が必要である。 非分裂細胞は、それぞれが中心体と呼ばれる一つのMTOCを持っています。 (これは上のビデオにも描かれています)。 核の近くに位置し、中心体は(-)端が尖っており、(+)端が細胞周辺を指している微小管の半径方向の構成にハブである。
中心体は、中心小体と呼ばれる2本の円柱で構成されています。 それぞれの中心小体は、9組の3つの横方向に融合した微小管からなり、核形成を促進するもの、特にγ–チューブリン環複合体(γチューブリンはTUBG_遺伝子によってコードされている)が豊富な非晶質の”中心小体周囲物質”に囲まれている。 γ-tubulinは”割れ目の洗濯機”としてについて考えられます:

モデルは、分割された末端が、γ−チューブリンがα−チューブリンに結合することを可能にすることである−すなわち、形成される微小管の(−)末端−が、微小管が形成されるための核形成種子を提供する。
微小管はin vitroで形成することができる。 動力学は主に臨界濃度に依存する。 (-)末端は(+)末端よりも重合に向かって傾斜していないので、より高い臨界濃度を有する。 実際の濃度が両端の臨界濃度の間にある場合、treadmillingは発生しません。
微小管が突然解離し始めるとき、これを「大惨事」と呼びます。 大災害には興味深いエネルギッシュなダイナミクスがあります。 伸長が起こる(+)末端であるβ-チューブリンは、GTP結合またはGDP結合のいずれかであり得ることを以前から思い出してください。 それはgtp結合しているとき、その微小管のより安定な部分であり、GDP結合チューブリンは解離しがちである。 微小管は、主に(+)末端にGTP結合β−チューブリンを添加することによって形成されるが、添加された後、β−チューブリン分子は後にGTPを加水分解し、GDP結合を残す。 そのため、gtp結合した微小管の一種の「先端」があり、より長い前に追加された微小管の下のより深いベータチューブリンはGDP結合しています。 GTP加水分解のフロンティアが先端に追いつく場合、または何かが微小管を切断するために起こる場合、その後、安定性の低い、GDP結合βチューブリンが露出 これが起これば、微小管はわらの下のどこかでgtp結合したベータtubulinの”島”に当るまで分解します。 大惨事は、末端をキャップし、プロトフィラメントを安定化させ、微小管が再形成することを可能にする新しいGTP結合チューブリン二量体の添加によって”救出”することができる。 このビデオでは、微小管の解重合が重合よりもはるかに高速であることがわかります:
微小管を研究するのに役立ついくつかの化合物があります。 抗痛風薬であるコルヒチンは、遊離α-β二量体に結合し、微小管形成のためのそれらの供給を減少させ、したがって解重合を促進する。 ノコダゾールはまた、新しい微小管の形成を妨害し、新しい微小管の形成が有糸分裂にとって重要であるため、ノコダゾールは抗腫瘍性癌薬である。 逆に、もう一つの抗がん剤であるパクリタキセル(タキソール)は、既存の微小管の分解も有糸分裂にとって重要であるため、微小管を安定化させることによ
微小管関連タンパク質(MAPs)は様々な役割を持っています。 注目すべきは、MAP4(非神経細胞)、MAP2(ニューロン)およびTau(ニューロン;MAPT遺伝子によってコードされる)である。 これらのすべては、それらが微小管を安定化させるように作用し、微小管の成長を支持し、大惨事に対して速度論をシフトさせることを共通している。 それぞれは微小管の否定的な部分に結合する正に荷電したアミノ酸が付いている18アミノ酸の伸張を含んでいます。 それらは、複数の微小管を一緒に束ねるように作用し、MAP2は、その長い腕(タウと比較して)のために、より大きな距離で微小管を保持する。
マップはリン酸化によって調節されています: MARKプロテインキナーゼは、MAPタンパク質中のs、TまたはYアミノ酸にリン酸基を共有結合し、微小管に結合する能力を低下させる。 (サイクリン依存性キナーゼはまた、有糸分裂の間にMAPsを調節する)。 これらのマップは、細胞の物理的構造を決定するために作用すると考えられている。 神経細胞では、MAP2は樹状突起に見られ、タウは主に軸索にある。 タウをコードするMAPT遺伝子の変異は、前頭側頭性認知症(FTD)を引き起こす。 過リン酸化タウは、アルツハイマー病の患者の脳に(理由はわかりませんが)見られます。 これらの疾患の両方のマウスモデルは、軸索変性を示し、タウは軸索の微小管を安定化させる仕事をすることができないことと一致する。
微小管の(+)末端への結合のために+Tipsという名前の別のクラスのタンパク質は、大惨事から保護することができます。 彼らは微小管に公正な方法を拡張するように見えます。 このビデオでは、微小管の末端が成長しているためにeb-1が外側に押されていることを示しています:
他の末端結合タンパク質は、安定性ではなく破局 キネシン-13はプロトフィラメントの終わりを湾曲させ、大惨事の閾値を低下させる働きをする。 Stathmin(STMN1遺伝子;別名Op18、Opはoncoproteinの略)は、即時のカタストロフィーとおそらくGTP加水分解の両方を促進し、プロトフィラメント内のチューブリン二量体に結合し、gtp結合ベータチューブリンの”島”を除去することにより、カタストロフィーを支持する長期的な投資の多くを表すだろう。 スタチミンはリン酸化によって調節される。 刀人(かたにん)は、文字通り微小管を切断する。
微小管モータータンパク質は、主にマイクロフィラメントモータータンパク質に類似しています。 それらは2つの家族入って来:(+)端の方にanterogradeすなわち動くkinesins;および(-)端の方に逆行すなわち動くほとんどがdyneins。 彼らはこのように”歩く”:
キネシンとダイニンは、移動するオルガネラ、エンドサイトーシスとエキソサイトーシス、減数分裂&有糸分裂中の染色体分離に関
キネシンの中で最もよく研究されているキネシン-1は、二つの重鎖単位(KIF1遺伝子など)で構成される四量体として作用するという点で”従来の”キネシンである。 ATPを加水分解して微小管に結合する「頭部」ドメインを含む2つの軽鎖単位(KLC遺伝子、例えばKLC1)、および貨物(この場合は小胞)に結合する「尾部」を含む2つの軽鎖単位(KLC遺伝子、例えばklc1)が含まれる。 リンカードメイン(この部分はどのタンパク質ですか?)は、二つの重鎖の二量体化を可能にする。
ここでは、アクションでそれのビデオです。 ビデオはそれを四量体ではなく二量体と呼んでいることに注意してください。
他のキネシンのいくつかは少し異なります:
- キネシン-2は、小胞&オルガネラ輸送も行うが、2つの異なる(関連しているが)重鎖とその貨物を調節するために使用する別のポリペプチドを有するヘテロトリマーである。
- キネシン-5は頭と尾の代わりに両端に頭を持っているので、2つの異なる微小管上で反対方向に歩き、それらを一緒に引っ張っています。 これは”バイポーラ運動”と呼ばれています。
- キネシン-14は、(-)末端に向かって移動する唯一の既知のキネシンであり、有糸分裂に関与している。
“歩く”運動の原理は、すべてのキネシンでほぼ同じであり、上記の最後のビデオに描かれているものです。 微小管に結合していない場合、「頭部」は両方ともADP結合である。 一つの頭部は微小管に遭遇し、それに結合し、そのADPを放出し、ATPがそれを置き換えることを可能にする。 ATPの結合はmicrotubuleに結合する指導的な位置に他の、”遅れ”の頭部を先に揺するリンカーで引っ張るconformational変更を引き起こします。 元のヘッドは、リン酸塩(ビデオではPiと略されています)を放出するATPを加水分解し、微小管から壊れているこのプロセスのエネルギー的に上り坂の
ミオシン(マイクロフィラメントを歩くモータータンパク質)とキネシンは非常に類似した構造を持っていますが、アミノ酸配列の類似性はありません。 したがって、それらはパラログではなく、むしろ収束進化の例であると考えられている。
人々はキネシンについて部分的に多くのことを話しています。 ダイニンは巨大なタンパク質で、2つの大きなサブユニット、2つの中間ユニット、2つの小さなサブユニットからなる。 彼らの膨大なサイズは、それらを分離して特徴付けることを困難にし、その働きはよく理解されていません。 私たちは、ダイナクチン複合体(マルチタンパク質複合体;ダイナクチン自体はDTCN_遺伝子である)が、ダイニンを貨物にリンクする”アダプター”として関与していることを知っている。
このビデオは、先週とこの講義からの材料の多くで全体の細胞骨格とロープをまとめたものです:
prp
分泌経路ノートに記載されているように、PrPは膜にGPIアンカーされ、非常に定期的にエンドサイトーシスを受け、PrPを含むエンドサイトーシス小胞を作 Encalada2011は、キネシン-1CおよびDHC1(ダイニン重鎖1)は、それぞれ前向きおよび逆行性これらのPrP小胞を輸送するための責任があることを見出します。 エンカラダがノックアウト、ノックダウン、またはキネシン-1Cを阻害すると、前行性および逆行性の動きが減少し、DHC1が破壊されたときと同様に減少した。 したがって、これらの2つの複合体は、反対方向に移動しますが、互いに活性化しているようです。 そして興味深いことに、これらのモータータンパク質を破壊することは、PrP小胞がモーターと関連するのを防ぐことはできませんでした。 この論文には、小胞がモータータンパク質をどのように活性化するか、および輸送の方向がどのように決定されるかについての他の多くの細胞生物学