세포 생물학 06:세포 골격 제 2 부:튜 불린
하버드 익스텐션의 세포 생물학 과정 강의 6 의 노트입니다.
지난 주 우리는 액틴으로 만들어진 마이크로 필라멘트를 다루었습니다. 이번 주:튜 불린으로 만든 미세 소관. 왜 세포 골격에는 두 개의 별도 시스템이 필요합니까? 마이크로 필라멘트는 개별 하위 단위의 사슬 일 뿐인 반면,미세 소관은 말 그대로 튜브(곧 볼 수 있듯이)–이량 체로 만든 체인으로 만든 벽을 가진 중공 실린더. 이러한 서로 다른 구조는 서로 다른 기능을 의미합니다: 마이크로 필라멘트는 자발적으로 더 쉽게 형성 될 수 있으며,서로 다른 네트워크 모양을 형성하고 셀의 다른 부분을 연결하기 위해(아프 2/3 및 지난 번에 논의 된 다른 복합체의 도움으로)분기 할 수 있습니다. 미세 소관은 형성하기 위해 핵 생성에 더 많이 의존하고,기본적으로 세포의 주변에 중심체를 연결하는 스포크 네트워크입니다. 실제로,마이크로 필라멘트는 세포 피질에 풍부하고 수축 운동 및 세포 운동성에 크게 관여하는 반면,미세 소관은 세포 기관을 조직하고 전 행성 및 역행 수송을위한 트랙 역할을하는 데 가장 관여합니다.
미세 소관
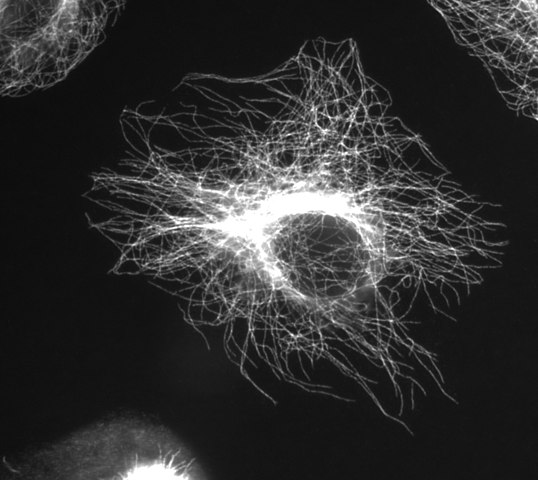
(위키미디어 공용 사용자 제프리 81 에 대한 이미지 감사합니다)
위키백과 외에도 이전 버전의 로디쉬와 쿠퍼 세포 생물학 교과서 또한 훌륭한 참고 자료입니다.
미세 소관의 관형 구조는 마이크로 필라멘트보다 견고하여 많은 중장비 당기기 및 밀기를 허용합니다. 그러나,미세소관은 온도에 따라 달라지므로,4 로 냉각되면 탈중합되고,37 로 가열되면 다시 재중합된다.
미세 소관의 기본 빌딩 블록은 튜빌린/튜빌린-튜불린 이량 체(각각 튜바_유전자 및 튜비_유전자에 의해 인코딩 됨)이다. 각각의 튜 불린 단백질은 각각~55 킬로다,그래서~110 다/아미노산 500 아미노산의 순서에. 모두 α-β-tubulin 바인딩 GTP 지만,α 거의 그냥 보유하고 그것에 영원히 반면에 β 그것을 교환할 수 있는 가수분해 GDP 다음으로 교환을 위한 새로운 GTP again.
연속된 스트랜드/스트랜드 이량 체는 프로토필라멘트를 구성하고,실린더에 나란히 배열된 13 개의 프로토필라멘트는 미세소관을 만든다. 프로토 필라멘트 사이의 공간을’솔기’라고합니다. 전체 미세 소관은 직경이~25 나노입니다. 당신은 미세 소관에 대한 세포 세그먼트의 내부 생활에서 그 구조(이량 체와 프로토 필라멘트로 만들어진)를 볼 수 있습니다:
각 이량체 내에서,상기 제 2 부단위는 중합을 위해 선호되는(+)말단이고,상기 제 2 부단위는 해중합을 위해 선호되는(-)말단이다. 미세 소관은 기본적으로 핵 형성,신장 및 정상 상태와 같은 미세 필라멘트와 동일한 3 단계로 형성됩니다. 그러나 마이크로 필라멘트와는 달리 그들은 스스로 쉽게 핵 생성하지 않으므로 핵 생성에는 미세 소관 조직 센터가 필요합니다. 비분열 세포는 각각 중심체라고 불리는 하나의 기동특무부대를 가지고 있다. (이것은 또한 위의 비디오에 묘사되어 있습니다). 핵 근처에 위치한 중심체는(-)끝이 뾰족하고(+)끝이 세포 주변을 가리키는 미세 소관의 방사형 구성에 허브입니다.
중심체는 중심체라고 불리는 2 개의 실린더로 구성된다. 각 중심체는 3 개의 옆으로 융합 된 미세 소관으로 구성된 각 9 세트로 구성되며,핵 생성을 촉진하는 물질이 풍부한 비정질’과피 세포 물질’으로 둘러싸여 있습니다.특히 감마 튜 불린 고리 복합체(감마 튜 불린은 튜 불린_유전자에 의해 암호화됩니다). 튜 불린은’분할 와셔’로 생각됩니다.:

이 모델은 분할 단부가 형성 될 미세 소관의(-)끝-미세 소관이 형성 될 핵 형성 시드를 제공하는 분할 단부가 형성 될 미세 소관의(–)끝 부분에 결합 할 수 있도록하는 것입니다.
미세 소관은 시험관 내에서 형성 될 수있다. 역학은 주로 임계 농도에 달려 있습니다. (-)단부는(+)단부보다 중합쪽으로 덜 기울어지기 때문에 더 높은 임계 농도를 갖는다. 실제 농도가 두 끝의 임계 농도 사이에 있으면 트레드 밀링이 발생합니다.
미세 소관이 갑자기 해리되기 시작하면 이것을’재앙’이라고합니다. 재앙에는 흥미로운 에너지 역학이 있습니다. 베타-튜 불린,이는(+)신장이 발생하는 끝,둘 중 하나 일 수 있습니다. 그것은 그것의 미세 소관의 더 안정한 부분이다. 미세 소관은 주로(+)끝에 베타-튜 불린 결합의 첨가에 의해 형성되지만,첨가 된 후 베타-튜 불린 분자는 나중에 가수 분해됩니다. 베타-튜 불린은 더 오래 전에 추가 된 미세 소관의 더 깊은 곳에서 국내 총생산 바인딩됩니다. 만약 거대자극자극성 가수분해의 경계선이 끝부분에 닿거나,어떤 일이 일어나 미세소관을 절단한다면,덜 안정되고,국내생산 결합된 베타-튜 불린이 노출되고,프로토필라멘트는”숫양의 뿔”처럼 벗겨지기 시작할 것이다. 이 경우 미세 소관은 짚 아래 어딘가에 베타 튜 불린의”섬”에 닿을 때까지 분해됩니다. 대재앙은 끝 부분을 캡핑하고 프로토 필라멘트를 안정화시켜 미세 소관이 다시 형성 될 수 있도록하는 새로운 관 결합 튜 불린 이량 체를 추가하여’구출’될 수 있습니다. 이 비디오에서 미세 소관의 해중합이 중합보다 훨씬 빠를 수 있음을 볼 수 있습니다:
다음은 미세 소관을 연구하는 데 유용한 몇 가지 화합물입니다. 통풍 방지 약물 인 콜히친은 유리 알파-베타 이량 체를 결합하여 미세 소관 형성을위한 공급을 감소시켜 해중합을 촉진합니다. 노코 다졸은 또한 새로운 미세 소관 형성을 방해하며,새로운 미세 소관의 형성이 유사 분열에 중요하기 때문에 노코 다졸은 항 종양 성 암 약물입니다. 반대로,다른 항암제 인 파클리탁셀(탁솔)은 기존 미세 소관의 분해가 유사 분열에도 중요하기 때문에 미세 소관을 안정화시킴으로써 작동합니다.
미세 소관 관련 단백질(지도)은 다양한 역할을합니다. 주목할만한 것은 맵 4(비 뉴런 세포),맵 2(뉴런)및 타우(뉴런,맵 유전자에 의해 코딩 됨)입니다. 이 모든 것들은 공통적으로 미세 소관을 안정화시키는 역할을 하며,미세 소관의 성장과 재앙에 대항하는 동역학을 이동시킵니다. 각 미세 소관의 부정적인 부분에 결합하는 양전하를 띤 아미노산과 18 아미노산 스트레치가 포함되어 있습니다. 그들은 여러 개의 미세 소관을 함께 묶는 역할을 할 수 있으며,맵 2 는 더 긴 팔(타우에 비해)으로 인해 미세 소관을 더 먼 거리에 고정시킵니다.
지도는 인산화에 의해 조절된다.: 마크 단백질 키나아제는 인산염 그룹을 공유 결합 에스,티 또는 와이 아미노산 지도 단백질에서 미세 소관을 결합하는 능력을 감소시킵니다. (사이클린 의존성 키나아제는 또한 유사 분열 동안지도를 조절합니다). 이 지도는 세포의 물리적 구조를 결정하는 역할을한다고 생각됩니다. 뉴런에서 맵 2 는 수상 돌기에서 발견되며 타우는 대부분 축삭에 있습니다. 타우를 코딩하는 맵 유전자의 돌연변이는 전 측두엽 치매를 유발합니다. 과인산화 된 타우는 알츠하이머 환자의 뇌에서 발견됩니다(우리는 그 이유를 알지 못하지만). 이 두 질병의 마우스 모델은 축삭 변성을 보여 주며,타우는 축삭에서 미세 소관을 안정화시키는 일을 할 수 없습니다.
미세 소관의(+)끝에 결합하는+팁이라는 이름의 또 다른 종류의 단백질은 재앙으로부터 보호 할 수 있습니다. 그들은 미세 소관 아래로 공정한 방법을 확장 나타납니다. 다른 말단 결합 단백질은 안정성보다는 재앙을 촉진한다. 키네신-13 은 프로토 필라멘트의 끝을 곡선으로 만들어 재앙의 문턱을 낮 춥니 다. 이는 베타-튜 불린의”섬”을 제거함으로써 재앙에 유리한 장기 투자를 더 많이 나타낼 것입니다. 스타트 민은 인산화에 의해 조절됩니다. 카타닌(일본 검 카타나 이후)은 말 그대로 미세 소관을 절단합니다.
미세소관 모터 단백질은 미세필라멘트 모터 단백질과 크게 유사하다. 그들은 두 가족으로옵니다:키네신,대부분은 전계 즉(+)끝을 향해 이동;그리고 다인,대부분은 역행 즉(-)끝을 향해 이동합니다. 그들은 다음과 같이’걷는다’:
키네신과 다이네인은 감수 분열&유사 분열 동안 세포 기관,세포 내 세포 세포 및 세포 외 세포 세포 및 염색체 분리에 관여한다.
키네신 중에서 가장 잘 연구 된 키네신-1 은 두 개의 중쇄 단위로 구성된 사량체로서 작동한다는 점에서’통상적 인’키네신이다. 본 발명은 본 발명은 본 발명은 본 발명은 본 발명은 본 발명은 본 발명은 본 발명은 본 발명은 본 발명은 본 발명은 본 발명은 본 발명은 본 발명은 본 발명은 본 발명은 본 발명은 본 발명은 본 발명은 본 발명은 본 발명은 본 발명은 본 발명은 본 발명은 본 발명은 본 발명은 본 발명은 본 발명은 본 발명은 본 발명은 본 발명은 본 발명은 본 발명은 본 발명은 본 발명은 본 발명은 본 발명은 본 발명은 본 발명은 본 발명은 링커 도메인(이 부분은 어떤 단백질입니까?)두 개의 중쇄를 이량 체화 할 수 있습니다.
여기 행동에 대한 비디오가 있습니다. 비디오 대신 사량체의 이량 체를 호출 참고;나는 그들이 단지 머리를 고려하고 꼬리를 논의하지 않기 때문에 그 생각.
다른 키네신 중 일부는 약간 다릅니다.:
- 소포&세포 기관 수송을 수행하는 키네신-2 는 두 개의 서로 다른(관련이 있지만)중쇄와 그화물을 규제하는 데 사용하는 또 다른 폴리펩티드를 갖는 헤테로 트리머이다.
- 키네신-5 는 머리와 꼬리 대신 양쪽 끝에 머리가 있어서 서로 다른 두 개의 미세 소관에서 반대 방향으로 걸어가면서 서로 끌어당긴다. 이를’양극성 운동’이라고합니다.
- 키네신-14 는(-)끝으로 이동하는 유일한 알려진 키네신이며 유사 분열에 관여합니다.
‘걷기’운동의 원리는 모든 키네신에 대해 거의 동일하며 위의 마지막 비디오에서 묘사 된 것입니다. 미세 소관에 결합되지 않을 때,’머리’는 둘 다 결합된다. 한 머리가 미세 소관을 만나서 그 머리에 결합하여 그 유도체를 방출하여 그것을 대체 할 수 있습니다. 이 미세 소관에 결합 위치를 선도하는 위치로 앞으로 다른’지체’머리를 스윙 링커에 끌어 구조적 변화를 유도한다. 그 후,원래의 헤드는 인산염(영상에서 파이로 축약됨)을 방출하는 인산염을 가수분해하고,이 과정에서 정력적으로 오르막 단계를 위한 에너지를 제공합니다.
미오신(마이크로 필라멘트를 걷는 운동 단백질)과 키네신은 매우 유사한 구조를 가지고 있지만 아미노산 서열 유사성은 없습니다. 따라서 그들은 보조자가 아니라 수렴 진화의 예라고 믿어집니다.
사람들은 부분적으로 키네신에 대해 많이 이야기합니다. 다인은 2 개의 대형,2 개의 중간 및 2 개의 작은 서브 유닛으로 만들어진 거대한 단백질입니다. 그들의 깎아지른 듯한 크기 그들을 분리 하 고 특성화 하기 어려운 했다 그리고 그들의 동작 잘 이해 되지 않습니다. 우리는 다이나틴 복합체(다단백질 복합체;다이나틴 자체는 유전자다)가 다이나인을 화물과 연결하는’어댑터’로 관여한다는 것을 알고 있습니다.
이 비디오는 지난 주 및이 강의 자료의 많은 부분에서 전체 세포 골격과 밧줄을 요약 한 것입니다:분비 경로 노트에서 언급한 바와 같이,세포막 내피세포는 세포막에 고정되어 있으며 매우 규칙적으로 세포내피세포가 생성되어 세포내피세포와 함께 소포가 생성된다. 그 결과,이들 중쇄는 그 중쇄를 운반 할 수 있고,그 중쇄는 그 중쇄를 운반 할 수 있고,그 중쇄는 그 중쇄를 운반 할 수 있으며,그 중쇄는 그 중쇄를 운반 할 수 있으며,그 중쇄는 그 중쇄를 운반 할 수 있으며,그 중쇄는 그 중쇄를 운반 할 수 있으며,그 중쇄는 그 중쇄를 운반 할 수있다. 엔 칼라다가 기절,기절 또는 억제되었을 때 키네신 -1 기음,전방위 및 역행 운동이 감소되었고,유사하게 디딜 방아 1 이 중단되었을 때. 그래서이 두 복합체는 반대 방향으로 이동하지만 서로를 활성화시키는 것 같습니다. 그리고 흥미롭게도,이러한 운동 단백질을 교란시키는 것은 포낭의 소포가 모터와 연결되는 것을 막지 못했습니다–그들은 단지 빨리 또는 자주 움직이지 않았습니다. 이 논문은 소포가 모터 단백질을 활성화하는 방법과 수송 방향이 어떻게 결정되는지에 대한 여러 가지 다른 세포 생물학 결론을 가지고 있습니다.