Biologi For Majors i
Læringsutbytte
- Forstå forskjellen mellom pre-mRNA og mRNA
etter transkripsjon må eukaryote pre-mrna gjennomgå flere behandlingstrinn før de kan oversettes. Eukaryote (og prokaryote) trnaer og rrnaer gjennomgår også behandling før de kan fungere som komponenter i proteinsyntesemaskinen.
mRNA-Behandling
den eukaryotiske pre-mRNA gjennomgår omfattende behandling før den er klar til å bli oversatt. De ekstra trinnene som er involvert i eukaryotisk mRNA-modning, skaper et molekyl med en mye lengre halveringstid enn en prokaryotisk mRNA. Eukaryote mrna varer i flere timer, mens den typiske E. coli mRNA varer ikke mer enn fem sekunder.
Pre-mrna blir først belagt I RNA-stabiliserende proteiner; disse beskytter pre-mRNA fra nedbrytning mens den behandles og eksporteres ut av kjernen. De tre viktigste trinnene i pre-mRNA-behandling er tilsetning av stabiliserende og signalerende faktorer ved 5′ og 3 ‘ ender av molekylet, og fjerning av mellomliggende sekvenser som ikke spesifiserer de riktige aminosyrene. I sjeldne tilfeller kan mrna-transkripsjonen «redigeres» etter at den er transkribert.
5 ‘Capping
mens pre-mRNA fortsatt syntetiseres, legges en 7-metylguanosinhette til 5′ enden av det voksende transkripsjonen ved en fosfatbinding. Denne delen (funksjonell gruppe) beskytter den naserende mRNA fra nedbrytning. I tillegg gjenkjenner faktorer involvert i proteinsyntese hetten for å bidra til å initiere oversettelse av ribosomer.
3 ‘ Poly-A Tail
når forlengelsen Er fullført, spaltes pre-mRNA av en endonuklease mellom EN aauaaa konsensussekvens og EN GU-rik sekvens, slik AT AAUAAA sekvensen på pre-mRNA. Et enzym kalt poly-a polymerase legger deretter til en streng på omtrent 200 a-rester, kalt poly-a-halen. Denne modifikasjonen beskytter ytterligere pre-mRNA fra nedbrytning og signalerer eksporten av de cellulære faktorene som transkripsjonen trenger til cytoplasma.
Pre-mRNA Spleising
Eukaryote gener er sammensatt av eksoner, som tilsvarer proteinkodende sekvenser (ex-on betyr at de uttrykkes), og mellomliggende sekvenser kalt introner (intron betegner deres mellomliggende rolle), som kan være involvert i genregulering, men fjernes fra pre-mRNA under behandling. Intronsekvenser i mRNA koder ikke funksjonelle proteiner.
oppdagelsen av introner kom som en overraskelse for forskere på 1970-tallet som forventet at pre-mrna ville spesifisere proteinsekvenser uten videre behandling, som de hadde observert i prokaryoter. Gener av høyere eukaryoter inneholder ofte en eller flere introner. Disse regionene kan tilsvare regulatoriske sekvenser; imidlertid er den biologiske betydningen av å ha mange introner eller å ha svært lange introner i et gen uklart. Det er mulig at introner reduserer genuttrykk fordi det tar lengre tid å transkribere pre-mrna med mange introner. Alternativt kan introner være ikke-funksjonelle sekvensrester igjen fra fusjonen av gamle gener gjennom evolusjonen. Dette støttes av det faktum at separate eksoner ofte koder for separate proteinunderenheter eller domener. For det meste kan sekvensene av introner muteres uten å påvirke proteinproduktet i siste instans.
alle pre-mrnas introner må fjernes helt og nøyaktig før proteinsyntese. Hvis prosessen feiler med enda et enkelt nukleotid, vil leserammen for de gjenforente eksonene skifte, og det resulterende proteinet vil være dysfunksjonelt. Prosessen med å fjerne introner og koble til eksoner kalles spleising (Figur 1). Introner fjernes og degraderes mens pre-mRNA fortsatt er i kjernen. Spleising skjer ved en sekvensspesifikk mekanisme som sikrer at introner vil bli fjernet og eksoner gjenforenes med nøyaktigheten og presisjonen av et enkelt nukleotid. Spleising av pre-mrna utføres av komplekser av proteiner og RNA-molekyler kalt spliceosomer.
Øvingsspørsmål
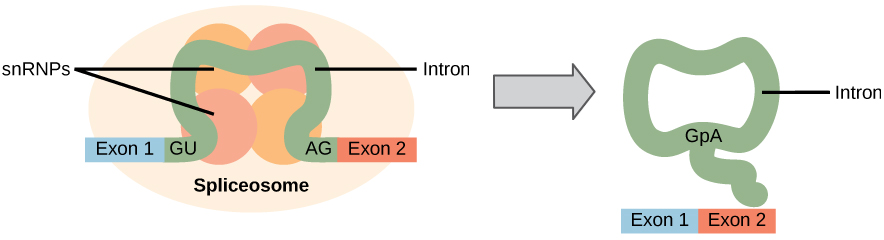
Figur 1. Pre-mRNA spleising innebærer nøyaktig fjerning av introner fra det primære RNA-transkripsjonen. Spleiseprosessen katalyseres av proteinkomplekser kalt spliceosomer som består av proteiner og RNA-molekyler kalt snrna. Spliceosomer gjenkjenner sekvenser ved 5′ og 3 ‘ enden av intronen.
Feil i spleising er involvert i kreft og andre menneskelige sykdommer. Hva slags mutasjoner kan føre til spleising feil?
Merk at mer enn 70 individuelle introner kan være til stede—og hver må gjennomgå prosessen med spleising – i tillegg til 5 ‘ capping og tilsetning av en poly-a hale-bare for å generere et enkelt, oversettbart mRNA-molekyl.
Rna-Redigering I Trypanosomer
Figur 2. Trypanosoma brucei er forårsaket av sovesyke hos mennesker. Mrna av dette patogenet må modifiseres ved tilsetning av nukleotider før proteinsyntese kan forekomme. (kreditt: endring av arbeid Av Torsten Ochsenreiter)
trypanosomer er en gruppe protozoer som inkluderer patogenet Trypanosoma brucei, som forårsaker sovesyke hos mennesker (Figur 2). Trypanosomer, og nesten alle andre eukaryoter, har organeller kalt mitokondrier som leverer cellen med kjemisk energi. Mitokondrier er organeller som uttrykker SITT EGET DNA og antas å være restene av et symbiotisk forhold mellom en eukaryot og en oppslukt prokaryot. Mitokondrielt DNA av trypanosomer viser et interessant unntak Fra Det Sentrale Dogmet: deres pre-mrna har ikke riktig informasjon for å spesifisere et funksjonelt protein. Vanligvis er dette fordi mRNA mangler flere u nukleotider. Cellen utfører et ekstra rna-behandlingstrinn kalt RNA-redigering for å rette opp dette.
Andre gener i mitokondriegenomet koder For 40-til 80-nukleotidguide Rna. Ett eller flere av disse molekylene interagerer ved komplementær baseparing med noen av nukleotidene i pre-mRNA-transkripsjonen. Guide-RNA har imidlertid flere nukleotider enn pre-mRNA har u nukleotider å binde med. I disse områdene slår guiden RNA ut. De 3 ‘ ender av guide Rna har en lang poly-u hale, og disse u baser er satt inn i områder av pre-mRNA transkripsjon ved hvilken guide Rna er loopet. Denne prosessen er helt mediert AV RNA-molekyler. Det vil si at guide Rna-snarere enn proteiner-tjener som katalysatorer I RNA-redigering.
rna-redigering er ikke bare et fenomen av trypanosomer. I mitokondrier av noen planter blir nesten alle pre-mRNAs redigert. RNA-redigering har også blitt identifisert hos pattedyr som rotter, kaniner og til og med mennesker. Hva kan være den evolusjonære årsaken til dette ekstra trinnet i pre-mRNA-behandling? En mulighet er at mitokondriene, som er rester av gamle prokaryoter, har en like gammel rna-basert metode for å regulere genuttrykk. Til støtte for denne hypotesen varierer endringer i pre-mrna avhengig av cellulære forhold. Selv om det er spekulativt, kan PROSESSEN MED rna-redigering være en holdover fra en primordial tid da RNA-molekyler, i stedet for proteiner, var ansvarlige for katalyserende reaksjoner.
Prøv Det
Bidra!
Forbedre denne sidenlær Mer