Biologi For Majors II
Læringsutbytte
- Forstå tidligere og nye klassifikasjonssystemer for virus
fordi de fleste virus sannsynligvis utviklet seg fra forskjellige forfedre, er de systematiske metodene som forskere har brukt til å klassifisere prokaryote og eukaryote celler ikke veldig nyttige. Hvis virus representerer «rester» av forskjellige organismer, er selv genomisk eller proteinanalyse ikke nyttig. Hvorfor?, Fordi virus har ingen felles genomisk sekvens som de alle deler. FOR eksempel er 16s rRNA-sekvensen som er så nyttig for å bygge prokaryote fylogenier, ikke til nytte for en skapning uten ribosomer! Biologer har brukt flere klassifikasjonssystemer tidligere. Virus ble først gruppert etter felles morfologi. Senere ble grupper av virus klassifisert etter typen nukleinsyre de inneholdt, DNA ELLER RNA, og om deres nukleinsyre var enkelt-eller dobbeltstrenget. Imidlertid grupperte disse tidligere klassifiseringsmetodene virus forskjellig, fordi de var basert på forskjellige sett med tegn på viruset. Den mest brukte klassifiseringsmetoden i Dag kalles Baltimore classification scheme, og er basert på hvordan messenger RNA (mRNA) genereres i hver bestemt type virus.
Tidligere Klassifikasjonssystemer
Virus inneholder bare noen få elementer som de kan klassifiseres med: virusgenomet, typen kapsid og konvoluttstrukturen for de innhyllede virusene. Alle disse elementene har tidligere blitt brukt til viral klassifisering(Tabell 1 Og Figur 1). Virale genomer kan variere i typen genetisk materiale (DNA ELLER RNA) og dets organisasjon (enkelt – eller dobbeltstrenget, lineært eller sirkulært, og segmentert eller ikke-segmentert). I noen virus er ytterligere proteiner som trengs for replikasjon, forbundet direkte med genomet eller inneholdt i viral kapsid.
| Tabell 1. Virusklassifisering Etter Genomstruktur og Kjerne | |
|---|---|
| Kjerneklassifiseringer | Eksempler |
| RNA | Rabiesvirus, retrovirus |
| DNA | Herpesvirus, koppevirus |
| Enkeltstrenget | Rabiesvirus, retrovirus |
| Dobbeltstrenget | Herpesvirus, koppevirus |
| Lineær | Rabiesvirus, retrovirus, herpesvirus, koppevirus |
| Rundskriv | Papillomavirus, mange bakteriofager |
| Ikke-segmentert: genomet består av et enkelt segment av genetisk materiale | Parainfluensavirus |
| Segmentert: genomet er delt inn i flere segmenter | Influensavirus |
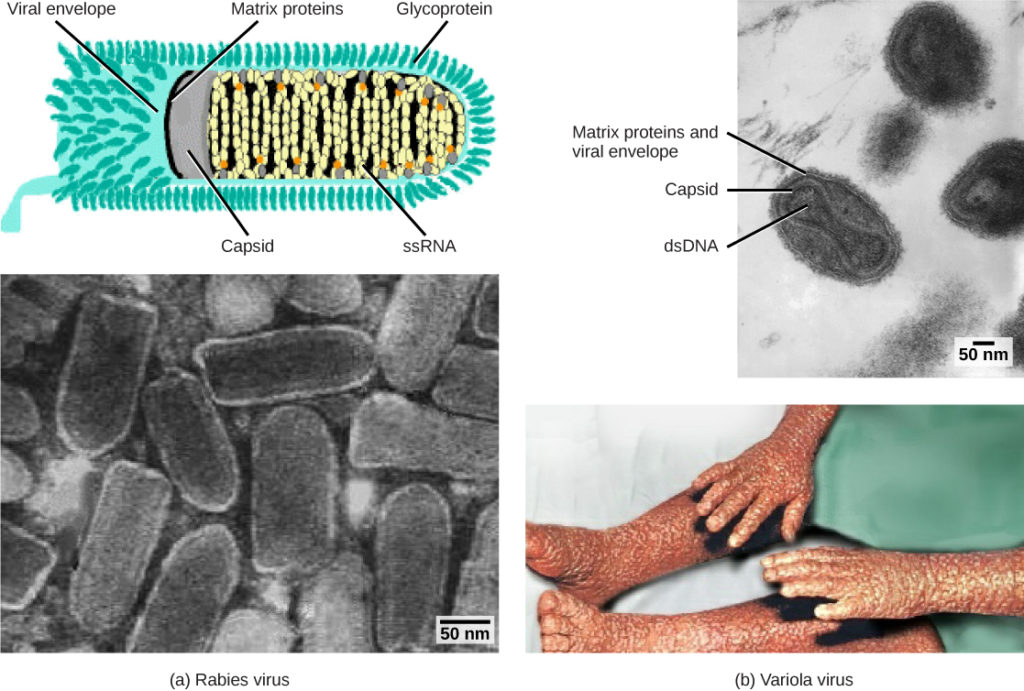
Figur 1. Virus er klassifisert basert på deres kjerne genetisk materiale og kapsiddesign. (A) Rabiesvirus har en enkeltstrenget RNA (ssRNA) kjerne og en innhyllet spiralformet kapsid, mens (b) variola-virus, det forårsakende middel for kopper, har en dobbeltstrenget DNA (dsDNA) kjerne og en kompleks kapsid. (credit «rabies diagram»: modifikasjon av ARBEID AV CDC; «rabies micrograph»: modifikasjon av arbeid Av Dr. Fred Murphy, CDC; credit «small pox micrograph»: modifikasjon av arbeid Av Dr. Fred Murphy, Sylvia Whitfield, CDC; credit «small pox photo»: modifikasjon av arbeid AV CDC; skala-bar data Fra Matt Russell)
Virus kan også klassifiseres etter utformingen av deres capsids(Tabell 2 Og Figur 2). Capsids er klassifisert som naken icosahedral, innhyllet icosahedral, innhyllet spiralformet, naken spiralformet og kompleks. Typen av genetisk materiale (DNA ELLER RNA) og dets struktur (enkelt – eller dobbeltstrenget, lineært eller sirkulært, og segmentert eller ikke-segmentert) brukes til å klassifisere virusets kjernestrukturer (Tabell 2).
| Tabell 2. Virusklassifisering Etter Kapsidstruktur | |
|---|---|
| Kapsidklassifisering | Eksempler |
| Naken icosahedral | Hepatitt a-virus, poliovirus |
| Innhyllet icosahedral | Epstein-Barr-virus, herpes simplex-virus, rubellavirus, gulfebervirus, HIV-1 |
| Innhyllet spiralformet | Influensavirus, kusma virus, meslinger virus, rabies virus |
| Naken spiralformet | Tobaksmosaikkvirus |
| kompleks med mange proteiner; noen har kombinasjoner av icosahedral og spiralformede kapsidstrukturer | Herpesvirus, koppevirus, hepatitt b-virus, t4 bakteriofag |

Figur 2. Transmisjonselektronmikrografer av ulike virus viser deres strukturer. Kapsid av (a) polio virus er naken icosahedral; (b) Epstein-Barr virus kapsid er innhyllet icosahedral; (c) kusma virus kapsid er en innhyllet helix; (d) tobakk mosaikk virus kapsid er naken spiralformet; og (e) herpesvirus kapsid er kompleks. (kreditt a: modifikasjon av arbeid Av Dr. Fred Murphy, Sylvia Whitfield; kreditt b: modifikasjon av arbeid Av Liza Gross; kreditt c: modifikasjon av arbeid Av Dr. Fa Murphy, CDC; kreditt d: modifikasjon AV arbeid AV USDA ARS; kreditt e: endring av arbeid Av Linda Stannard, Institutt For Medisinsk Mikrobiologi, University Of Cape Town, Sør-Afrika, NASA; skala-bar data Fra Matt Russell)
Baltimore Classification
det mest brukte systemet for virusklassifisering ble utviklet av Nobelprisvinnende biolog David Baltimore tidlig på 1970-tallet. I tillegg til forskjellene i morfologi og genetikk nevnt ovenfor, Grupperer Baltimore classification scheme virus i henhold til hvordan mRNA er produsert under virusets replikative syklus.
Gruppe i-virus inneholder dobbeltstrenget DNA (dsDNA) som deres genom. Deres mRNA er produsert ved transkripsjon på omtrent samme måte som med cellulært DNA.
Gruppe II-virus har enkeltstrenget DNA (ssDNA) som deres genom. De konverterer sine enkeltstrengede genomer til et dsdna-mellomprodukt før transkripsjon til mRNA kan forekomme.
Gruppe III-virus bruker dsRNA som deres genom. Strengene adskiller seg, og en av dem brukes som en mal for generering av mRNA ved BRUK AV RNA-avhengig rna-polymerase kodet av viruset.
Gruppe IV-virus har ssRNA som deres genom med positiv polaritet. Positiv polaritet betyr at det genomiske RNA kan tjene direkte som mRNA. Intermediater av dsRNA, kalt replikative mellomprodukter, er laget i ferd med å kopiere det genomiske RNA. Multiple, full-lengde rna tråder av negativ polaritet (gratis til den positive-strandet genomisk RNA) er dannet fra disse mellomprodukter, som deretter kan tjene som maler for produksjon AV RNA med positiv polaritet, inkludert både full lengde genomisk RNA og kortere viral mrna.
Gruppe V-virus inneholder ssRNA-genomer med negativ polaritet, noe som betyr at deres sekvens er komplementær til mRNA. Som Med Gruppe IV-virus brukes dsrna-mellomprodukter til å lage kopier av genomet og produsere mRNA. I dette tilfellet kan det negative strandede genomet konverteres direkte til mRNA. I tillegg er full lengde positive rna-tråder laget for å tjene som maler for produksjon av det negative-strandede genomet.
Gruppe VI-virus har diploide (to kopier) ssrna-genomer som må konverteres, ved hjelp av enzymet revers transkriptase, til dsDNA; dsDNA blir deretter transportert til kjernen i vertscellen og satt inn i vertsgenomet. Deretter kan mRNA produseres ved transkripsjon av det virale DNA som ble integrert i vertsgenomet.
Gruppe VII-virus har partielle dsdna-genomer og lager ssrna-mellomprodukter som fungerer som mRNA, men konverteres også tilbake til dsDNA-genomer ved revers transkriptase, som er nødvendig for genomreplikasjon. Egenskapene til hver gruppe I Baltimore klassifisering er oppsummert I Tabell 3 med eksempler på hver gruppe.
| Tabell 3. Baltimore Klassifisering | |||
|---|---|---|---|
| Gruppe | Egenskaper | Modus for mRNA-Produksjon | Eksempel |
| I | Dobbeltstrenget DNA | mRNA transkriberes direkte fra DNA-malen | Herpes simplex (herpesvirus) |
| II | Enkeltstrenget DNA | DNA omdannes til dobbeltstrenget form før RNA transkriberes | Hundeparvovirus (parvovirus) |
| III | Dobbeltstrenget RNA | mRNA er transkribert fra RNA-genomet | Gastroenteritt Hos Barn (rotavirus) |
| IV | Enkeltstrenget RNA (+) | Genomfunksjoner som mRNA | Forkjølelse (pircornavirus) |
| V | Enkeltstrenget RNA ( – ) | mRNA transkriberes fra rna-genomet | Rabies (rhabdovirus) |
| VI | enkeltstrengede RNA-virus med revers transkriptase | Revers transkriptase gjør DNA FRA RNA-genomet; DNA blir deretter innlemmet I vertsgenomet; mRNA transkriberes fra det inkorporerte DNA | Humant immunsviktvirus (HIV) |
| VII | Dobbeltstrenget DNA-virus med revers transkriptase | det virale genomet er dobbeltstrenget DNA, men viralt DNA replikeres gjennom et rna-mellomprodukt; RNA kan tjene direkte som mRNA eller som en mal for å lage mRNA | Hepatitt b-virus (hepadnavirus) |