Cellebiologi 06: Cytoskelettet Del II: Tubulin
dette er notater fra forelesning 6 Av Harvard Extension ‘ S Cellebiologi kurs.
I forrige uke dekket vi mikrofilamenter, som er laget av aktin. Denne uken: mikrotubuli, laget av tubulin. Hvorfor spør du, trenger cytoskelettet to separate systemer? Tenk på at mikrofilamenter bare er kjeder av individuelle underenheter, mens mikrotubuli er bokstavelig talt rør – som vi snart ser) – hule sylindere med vegger laget av kjeder laget av dimere. Disse forskjellige strukturer betyr forskjellige evner: mikrofilamenter kan lettere dannes spontant, og kan forgrene seg (Ved Hjelp Av Arp2/3 og andre komplekser diskutert forrige gang) for å danne forskjellige nettverksformer og koble til ulike deler av cellen. Mikrotubuli stole mer tungt på nukleasjon for å danne, og er i utgangspunktet et eikenettverk som forbinder sentrosomet til cellens periferi. I praksis er mikrofilamenter rikelig i cellebarken og tungt involvert i kontraktile bevegelser og cellemotilitet, mens mikrotubuli er mest involvert i å organisere organeller og tjene som spor for anterograd og retrograd transport.
Mikrotubuli
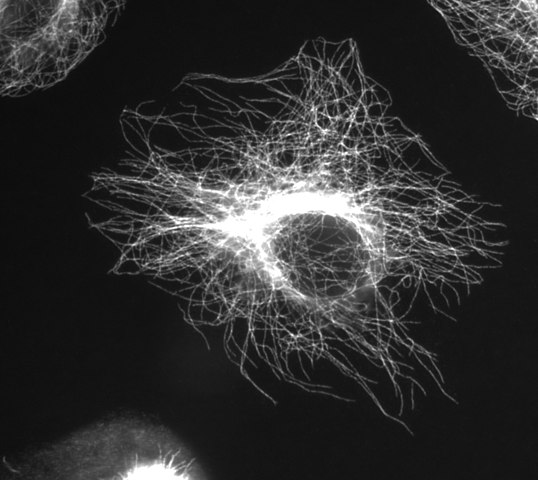
(Bilde takket Være Wikimedia Commons bruker Jeffrey81)
i Tillegg Til Wikipedia, tidligere utgaver Av Lodish og Cooper cellebiologi lærebøker tilgjengelig PÅ NCBI er også gode referanser.
den rørformede strukturen til mikrotubuli er kraftigere enn mikrofilamenter, noe som gir mye kraftig trekk og trykk. Selv om mikrotubuli er robuste, er de temperaturavhengige-de depolymeriserer hvis de avkjøles til 4°C og vil repolymerisere igjen hvis de oppvarmes til 37°C forutsatt AT GTP er tilgjengelig.
den grunnleggende byggesteinen til mikrotubuli er en α-tubuilin/β-tubulin dimer (kodet av HENHOLDSVIS TUBA_ gener og TUBB_ gener). De enkelte tubulinproteinene veier hver ~55kda, så ved ~110da / aminosyre som er i størrelsesorden 500 aminosyrer. Både α-og β-tubulin binder GTP, men α holder ganske mye bare på DET for alltid, mens i β kan det byttes eller hydrolyseres TIL BNP og deretter byttes mot ny GTP igjen.
Strenger av sammenhengende α / β dimerer utgjør protofilamenter, og 13 protofilamenter arrangert side om side i en sylinder gjør en mikrotubule. Mellomrommet mellom protofilamentene kalles en søm. Hele mikrotubulen er ~25 nm i diameter. Du kan se sin struktur (laget av dimerer og protofilamenter) I Det Indre livet Til Cellesegmentet om mikrotubuli:
innenfor hver dimer er β-underenheten ( + ) enden, favorisert for polymerisering og α er ( – ) enden, favorisert for depolymerisering. Mikrotubuli danner i utgangspunktet de samme tre trinnene som mikrofilamenter – nukleasjon, forlengelse og steady state. Men i motsetning til mikrofilamenter de ikke nucleate lett på egen hånd, så nucleation krever microtubule organisere sentre (MTOCs). Ikke-delende celler har hver BARE EN MTOC, kalt sentrosomet. (Dette er også avbildet i videoen ovenfor). Ligger nær kjernen, er sentrosomet hub til en radial konfigurering av mikrotubuli med ( – ) ender pekte inn og ( + ) ender peker mot cellens periferi.
sentrosomet består av 2 sylindere kalt sentrioler. Hver centriol består av hver 9 sett med 3 lateralt smeltede mikrotubuli, omgitt av et amorft ‘pericentriolært materiale’ rik på ting som fremmer nukleasjon – spesielt γ-tubulin ringkomplekser (gamma tubulin er kodet AV TUBG_-gener). γ-tubulin er tenkt som en ‘delt vaskemaskin’:

modellen er at de splittede endene tillater γ-tubulin å binde seg til α-tubulin – dvs. ( – ) enden av en å-være-formet mikrotubule – som gir kjernefrøet for en mikrotubule å danne.
Mikrotubuli kan dannes in vitro. Dynamikken avhenger hovedsakelig av kritiske konsentrasjoner. (- ) Enden er mindre tilbøyelig til polymerisering enn ( + ) enden, så den har en høyere kritisk konsentrasjon. Hvis den faktiske konsentrasjonen er mellom de to ender kritiske konsentrasjoner, tredemølle forekommer.
når en mikrotubule plutselig begynner å dissosiere, kalles dette en katastrofe. Katastrofer har noen interessante energiske dynamikker. Husk fra tidligere at beta-tubulin, som er (+) enden der forlengelse skjer, kan enten VÆRE GTP – eller BNP-bundet. Det er en mer stabil del av sin mikrotubule når DEN ER GTP-bundet; BNP-bundet tubulin er egnet til å dissociere. Mikrotubuli danner hovedsakelig VED tilsetning AV GTP-bundet beta-tubulin ved ( + ) enden, men etter å ha blitt tilsatt, hydrolyserer beta-tubulinmolekylene DERES GTP, og etterlater DEM BNP-bundet. Så det er en slags ‘tips’ av mikrotubulen SOM ER GTP-bundet, mens beta-tubulin dypere ned i mikrotubulen, lagt til lenger siden, ER BNP-bundet. Hvis grensen TIL GTP-hydrolyse fanger opp til spissen, eller hvis noe skjer for å kutte mikrotubulen, blir den mindre stabile, BNP-bundne beta-tubulin utsatt, og protofilamentene vil begynne å skrelle bort som «rams horn». Hvis dette skjer, vil mikrotubulen demontere til den treffer en » øy » AV GTP-bundet beta-tubulin et sted lenger ned i halmen. Katastrofe kan ‘reddes’ ved tillegg av nye gtp-bundet tubulin dimerer som vil dekke endene og stabilisere protofilamenter, slik at mikrotubuli å re-form. Du kan se i denne videoen at depolymerisering av mikrotubuli kan være mye raskere enn polymerisering:
her er noen forbindelser som er nyttige for å studere mikrotubuli. Colchicin, et anti-giktmedikament, binder frie alfa-beta-dimerer, reduserer forsyningen for mikrotubuldannelse og dermed fremmer depolymerisering. Nocodazol forstyrrer også ny mikrotubuldannelse, og fordi dannelse av nye mikrotubuli er viktig for mitose, er nocodazol et antineoplastisk kreftmedisin. Omvendt virker paclitaxel (taxol), et annet anti-kreft stoff, ved å stabilisere mikrotubuli, siden nedbrytning av eksisterende mikrotubuli også er viktig for mitose.
Mikrotubuliassosierte proteiner (Kart) har ulike roller. Bemerkelsesverdig ER MAP4 (i ikke-neuronale celler), MAP2 (i nevroner) og Tau (i nevroner; kodet AV MAPT-genet). Alle disse har til felles at de virker for å stabilisere mikrotubuli, skifte kinetikken til fordel for mikrotubulevekst og mot katastrofe. Hver inneholder en 18 aminosyrestrekning med positivt ladede aminosyrer som binder seg til den negative delen av mikrotubuli. DE kan fungere for å pakke flere mikrotubuli sammen , MED MAP2 som holder mikrotubuli i større avstand på grunn av sin lengre arm (sammenlignet med Tau).
Kartene er regulert av fosforylering: MARK-proteinkinasene binder kovalent fosfatgrupper til S -, T-eller y-aminosyrer i MAP-proteinene, noe som reduserer deres evne til å binde mikrotubuli. (Syklinavhengige kinaser regulerer Også Kartene under mitose). Det antas at Disse Kartene virker for å bestemme cellens fysiske struktur. I nevroner FINNES MAP2 i dendriter og Tau er for det meste i axonen. Mutasjoner i MAPT-genet som koder For Tau forårsaker frontotemporal demens (FTD). Hyperfosforylert Tau er funnet (selv om vi ikke vet hvorfor) I Alzheimers pasienters hjerner. Musemodeller av begge disse sykdommene viser aksonal degenerasjon, i samsvar med Tau ikke å kunne gjøre jobben sin med å stabilisere mikrotubuli i axonen.
En annen klasse proteiner, kalt +Tips for binding til (+) enden av mikrotubuli, kan beskytte mot katastrofe. De ser ut til å utvide en rettferdig måte ned i mikrotubulen. DENNE videoen viser AT EB-1 blir presset utover fordi endene av mikrotubuli vokser:
Andre endebindende proteiner fremmer katastrofe i stedet for stabilitet. Kinesin-13 virker for å kurve enden av protofilamentene, senke terskelen for katastrofe. Stathmin (STMN1 gen; aka Op18, Hvor Op står for oncoprotein) binder seg til tubulin dimerer innenfor et protofilament, fremme både umiddelbar katastrofe og muligens OGSÅ gtp hydrolyse, som ved å fjerne «øyer» AV GTP-bundet beta-tubulin ville representere mer av en langsiktig investering i favør av katastrofe. Stathmin reguleres ved fosforylering. Katanin (Etter Det Japanske sverdet katana) kutter bokstavelig talt mikrotubuli.
Mikrotubuli motor proteiner er i stor grad lik mikrofilament motor proteiner. De kommer i to familier: kinesiner, hvorav de fleste beveger anterograd dvs. mot ( + ) enden; og dyneins, hvorav de fleste beveger seg retrograd dvs. mot ( – ) enden. De’ gå ‘ som dette:
Kinesin og dynein er involvert i å flytte organeller, endocytose og eksocytose, og kromosom segregering under meiose & mitose.
Kinesin-1, den best studerte av kinesinene, er et konvensjonelt kinesin ved at det fungerer som en tetramer bestående av to tunge kjedeenheter (kif1-gener, f. eks. KIF1A) som utgjør hoveddomenene som hydrolyserer ATP og binder mikrotubuli, og to lette kjedeenheter (KLC-gener f. EKS. KLC1) som utgjør en hale som binder lasten, i dette tilfellet vesikler. Et linker domene (hvilket protein er denne delen av?) tillater dimerisering av to tunge kjeder.
her er en video av det i aksjon. Merk at videoen kaller det en dimer i stedet for en tetramer; jeg tror det er fordi de bare vurderer hodene og ikke diskuterer haler.
noen av de andre kinesinene er litt forskjellige:
- Kinesin-2, som også gjør vesikkel & organelltransport, er en heterotrimer med to forskjellige (skjønt relaterte) tunge kjeder og et annet polypeptid det bruker til å regulere lasten.
- Kinesin-5 har hoder i begge ender i stedet for hode og hale, slik at den går i motsatte retninger på to forskjellige mikrotubuli, og trekker dem sammen. Dette kalles bipolar bevegelse.
- Kinesin-14 er det eneste kjente kinesin som beveger seg mot ( – ) enden, og det er involvert i mitose.
prinsippene for ‘walking’ – bevegelsen er ganske mye det samme for alle kinesins, skjønt, og er det som er avbildet i den siste videoen ovenfor. Når de ikke er bundet til en mikrotubule, er hodene BEGGE ADP-bundet. Ett hode vil skje for å møte en mikrotubule og binde seg til DET, slippe ADP, slik AT EN ATP kan erstatte den. ATP-bindingen induserer konformasjonsendring som trekker på linkeren, svinger den andre,’ legger ‘ hodet fremover i ledende posisjon der det binder seg til mikrotubulen. Det opprinnelige hodet hydrolyserer DERETTER ATP som frigjør fosfat (som forkortes Som Pi i videoen) og gir energien til det ene energisk oppoverbakke trinnet i denne prosessen, som bryter bort fra mikrotubulen.
Myosin (motorproteinet som går på mikrofilamenter) og kinesin har svært like strukturer, men ingen aminosyresekvenslikhet. Dermed antas de ikke å være paraloger, men heller et eksempel på konvergent evolusjon.
Folk snakker mye om kinesiner delvis fordi vi ikke forstår dyneins ganske så godt. Dyneins er store proteiner, laget av 2 store, 2 mellomliggende og 2 små underenheter. Deres størrelse har gjort dem vanskelig å isolere og karakterisere og deres arbeid er ikke godt forstått. Vi vet at dynactin-komplekset (et multi-proteinkompleks; dynaktinene selv er dtcn_-genene) er involvert som en adapter som knytter dynein til last.
denne videoen oppsummerer hele cytoskjelettet og tauene i mye av materialet fra forrige uke og dette foredraget:
PrP
Som nevnt i sekretoriske banenotater, er PrP gpi-forankret til membranen og gjennomgår endocytose veldig regelmessig, og skaper endocytiske vesikler med PrP i dem. Encalada 2011 finner At Kinesin-1C OG DHC1 (dynein heavy chain 1) er ansvarlig for transport Av Disse PrP vesikler anterograd og retrograd henholdsvis. Når Encalada slått ut, slått ned eller hemmet Kinesin-1C, ble anterograd og retrograd bevegelse redusert, og på samme måte når DHC1 ble forstyrret. Så det ser ut til at disse to kompleksene, selv om de beveger seg i motsatte retninger, aktiverer hverandre. Og interessant, forstyrre disse motorproteinene forhindret Ikke PrP vesikler fra å knytte seg til motorer – de beveget seg ikke så fort eller så ofte. Papiret har en rekke andre cellebiologiske konklusjoner om arten av hvordan vesikler aktiverer motorproteiner og hvordan transportretningen bestemmes.