Grenser I Mikrobiologi
Introduksjon
Astroviridae-familien inneholder ikke-innhyllede, positive sans, enkeltstrengede RNA-virus innen to slekter, Mamastrovirus og avastrovirus, som infiserer henholdsvis pattedyr og fugler. For Tiden anerkjenner Den Internasjonale Komiteen For Taksonomi Av Virus (International Committee on Taxonomy Of Viruses , 2018) 19 arter, nemlig Mamastrovirus-1 til -19, innenfor Mamastrovirus-slekten; det er imidlertid mange stammer som venter på klassifisering, hvorav noen er foreløpig ansett som nye arter (Donato Og Vijaykrishna, 2017).
siden 2010 har flere astroviruser i økende grad blitt anerkjent som nevroinvasive i ulike pattedyrarter, inkludert mennesker (Quan et al., 2010; Naccache et al., 2015), mink (Blomstr@m et al., 2010), kveg (Li Et al., 2013), sau (Pfaff et al., 2017), og griser (Boros et al., 2017). Etter første anerkjennelse av bovin astrovirus-assosiert encefalitt I usa storfe (Li et al., 2013), viste en retrospektiv studie i tilfeller av sporadisk bovin encefalitt av ubestemt etiologi fra Sveits at dette neuroinvasive astroviruset hadde gått uoppdaget i flere tiår (Selimovic-Hamza et al., 2016). Selv om epidemiologi og overføringsruter for disse astrovirusene er ukjente, har overføring av kryssarter blitt foreslått basert på det høye identitetsnivået (> 98%), delt mellom bovin og sau neuroinvasive astrovirus ved nukleotid og aminosyre nivåer (Boujon et al ., 2017).
Bovine astrovirus (BoAstVs), kalt BoAstV-NeuroS1 (Li et al., 2013) Og BoAstV-CH13 (Bouzalas et al., 2014), ble opprinnelig funnet i hjernen av storfe med ikke-suppurativ encefalitt i henholdsvis usa og Sveits. Til tross for den forskjellige nomenklaturen representerer begge virusene samme genotype arter (Bouzalas et al., 2016; Selimovic-Hamza et al., 2017a) som fortsatt venter på offisiell klassifisering AV ICTV. I 2015 ble En tidligere ukjent BoAstV-stamme, Kalt BoAstV-CH15, identifisert i hjernen hos kyr med encefalitt I Sveits. Full genom fylogenetisk sammenligning viste et nærmere forhold Av BoAstV-CH15 med en sau astrovirus (OvAstV)enn Med BoAstV-CH13 (Seuberlich et al ., 2016). Koinfeksjon Med BoAstV-CH13 Og BoAstV-CH15 ble også dokumentert i ett tilfelle (Seuberlich et al., 2016). Samme år I Tyskland, Schlottau et al. (2016) rapporterte en roman astrovirus, nemlig BoAstV-BH89/14, i en ku med encefalitt, som var mest knyttet Til OvAstV og BoAstV-CH15. Deretter Ble BoAstV-CH13 / NeuroS1 identifisert i 2017 i tilfeller av bovin encefalitt i øst-Og vest-Canada (Spinato et al., 2017; Selimovic-Hamza et al.( 2017b). I 2018 ble en ny neuroinvasiv BoAstV nært beslektet Med Nordamerikanske Og Europeiske BoAstV-NeuroS1/BoAstV-CH13 identifisert i en steer med ikke-suppurativ encefalomyelitt I Japan, og forekomsten av intra-genotypisk rekombinasjon mellom De Nordamerikanske og Europeiske stammene ble foreslått (Hirashima et al., 2018).
mens tilfeller av astrovirus-assosiert encefalitt har blitt rapportert I Nord-Amerika, Europa og Asia, har deres tilstedeværelse aldri blitt dokumentert På Den Sørlige halvkule. Her beskriver vi et tilfelle av astrovirus-assosiert encefalitt hos storfe I Uruguay, som utvider den geografiske fordeling og genetisk mangfold av neuroinvasive astrovirus og gir fylogeografisk bevis som tyder på at dette viruset ble introdusert I Amerika Fra Europa og senere spredt Seg Til Asia.
Materialer Og Metoder
Historie Og Signalering
i juni 2018, en 22 måneder gammel Holstein styre i en gruppe på 37 styrer i en ∼300 hektar gård I Colonia, Uruguay, utviklet progressive nevrologiske synger inkludert uvanlig oppførsel, formålsløs gange, sirkle, ataksi, repeterende og ukoordinert tunge bevegelser, og recumbency. Flokken beite på en årlig havre beite og ble supplert med mais ensilasje. En presumptiv klinisk diagnose av cerebral listeriose av veterinæren førte til behandling med penicillin og streptomycin, men dyret døde spontant etter et klinisk kurs som varer 3 dager.
Patologisk Undersøkelse, In situ Hybridisering (ISH) og Immunhistokjemi (IHC)
styrehodet ble fjernet fra kadaveret og sendt TIL Inias Veterinary Diagnostic Laboratory (Animal Health Platform) for diagnostisk utredning. Halvparten av hjernen, et kort segment av proksimal cervikal ryggmargen (C1), trigeminal ganglion og rot av trigeminusnerven, spyttkjertel, retrofaryngeal lymfeknute, oropharynx, spiserør, tunge og skjelettmuskulatur, var nedsenkning-fast i 10% nøytral bufret formalin i 48-72 timer. Vev ble rutinemessig behandlet for histologi, innebygd i paraffin, mikrotom-seksjonert ved 4-5 µ og farget med hematoksylin Og eosin (H&E) Og Gram flekker.
Kromogen ISH ble utført manuelt på 5 µ seksjoner av formalin-fast, parafin-innebygd (FFPE) hjernestamme, cerebrum og cerebellum På Superfrost Plus-lysbilder (Thermo Fisher Scientific, Pittsburgh, PA, Usa) ved Bruk Av RNAscope 2.5 Red assay kit (Cat #322360, Advanced Cell Diagnostics, Hayward, CA, Usa) og BoAstV-sonden Cat. #406921. Sonden består AV 20ZZ par målretting region 5232-6180 av viruset (GenBank KF233994.1). Hver 5@m seksjon av vev ble forbehandlet med varme og protease før probe hybridisering i 2 timer ved 40°C og behandlet i henhold til produsentens anbefalinger. Negative kontroller som brukes for validering av signal inkludert en urelatert (GC-innhold matchet) sonde kjøre på serielle seksjoner og sondering vev fra uinfiserte dyr. Lysbilder ble counterstained med hematoxylin og montert Med EcoMount (Biocare Medical, Concord, CA, Usa).
I Tillegg BLE IHC utført I FFPE-seksjoner av hjernestamme, cerebrum og cerebellum, som tidligere beskrevet, for identifisering Av West Nile virus (WNV, Flavivirus) (Palmieri et al., 2011), rabiesvirus (Lyssavirus) (Stein Et al.( 2010), Og Chlamydia spp. (Giannitti et al., 2016) antigener.
Molekylær Virologi
Nukleinsyreutvinning ble utført fra en sammenslått prøve av frossen (-20°C) hjerne ved Bruk Av MagMAX Nukleinsyreisolasjonssett® (Thermo Fisher Scientific). For astrovirusdeteksjon ble reverstranskripsjon (RT) utført med Revertaid Reverstranskriptase® (Thermo Fisher Scientific) og tilfeldige heksamerprimere (Qiagen). PCR ble utført fra cDNA ved Bruk Av MangoMix® (Biolin)og primere som forsterker et 432-nukleotidfragment av astroviruspolymerasegenet (Tse et al., 2011). PCR-produktet ble visualisert i 2% agarosegel, renset Ved Bruk Av Purelink@ Quick Gel Extraction og PCR Purification Combo Kit (Invitrogen), og sekvensert Ved Macrogen Inc. (Seoul, Sør-Korea). For astrovirus hele genom amplifikasjon, Maxima H Minus Revers Transkriptase (Thermo Fisher Scientific)og oligo(dT) 18 for obtention av cDNA, Og MangoMix® (Biolin) eller Ranger DNA Polymerase (Biolin) med primere beskrevet Av Hirashima et al. 2018 ble brukt. PCR-produktene ble visualisert i 1-2% agarosegel, renset og sekvensert som nevnt ovenfor. Sekvensmontering ble utfort Med SeqMan (Lasergene 8, DNASTAR). Tjuefem komplette genomsekvenser av neuroinvasive astrovirus fra storfe, sau, griser, mennesker og mink og enterisk bovint astrovirus tilgjengelig I GenBank ble lastet ned og justert ved Hjelp Av Clustal W I MEGA 7-programvare (Kumar et al., 2016). W-IQ-TREE1 (Trifinopoulos et al., 2016) ble brukt til å bestemme den best egnede modellen for sekvensutvikling (SYM + I + G4) og å konstruere et maksimalt sannsynlighetsfylogenetisk tre med de nesten komplette sekvensene Av BoAstV oppdaget i dette tilfellet, og de komplette sekvensene lastet ned Fra GenBank, ved hjelp av bootstrap som statistisk metode for å vurdere clades robusthet. Likhet tomten ble utført Med SimPlot programvare (Lole et al., 1999). P-avstander på aminosyre nivå AV ORF2 ble estimert MED MEGA 7 programvare (Kumar et al., 2016).
I Tillegg ble En Bayesiansk fylogeografisk analyse utført med BEAST v1.8. 4-pakken (Drummond et al., 2012), ved hjelp av: Den komplette kodingsregionen Av BoAstV CH13 / NeuroS1 lineage, ORF1ab (ikke-strukturelle gener), ORF2 (strukturelt gen), ORF1a (protease) Og en delvis orf1b (polymerase genomisk region, For Hvilken Kanadiske stammer var tilgjengelige), med alle sekvensene tilgjengelige I GenBank (siste tiltredelse 18.April 2019), for å bestemme evolusjonær hastighet, alder/år av de vanlige forfedrene, og den mest sannsynlige ruten for viral sirkulasjon etter land (Sveits, Uruguay, Usa, Canada og Canada). japan). Mangel på rekombinasjon i datasettet ble bestemt Ved Hjelp Av Rekombinasjon Deteksjonsprogram 4. Substitusjonsmodellen som best passer til hver justering ble bestemt VED HJELP AV MEGA 7-programvare gjennom Bayesian information criterion (BIC) – verdier, og den tidsmessige strukturen til hvert datasett ble evaluert ved Hjelp Av TempEst (Rambaut et al., 2016). Lognormal avslappet molekylær klokke Med Bayesian Skyline analyse ble valgt Av Bayes Faktor blant de ulike kombinasjoner av molekylære klokker og coalescent tre priors brukes. Deteksjonslandet ble brukt som trekk. Markov kjeden Monte Carlo lengde var 100 millioner generasjoner, sikre konvergens av analysen, evaluert I Tracer v1. 6.0, og den bakre sannsynligheten ble brukt til å evaluere clades. Maksimal clade troverdighet treet (MCCT) ble oppnådd Ved Hjelp Av TreeAnnotator programvare FRA BEAST og visualisert I FigTree v1.4.3.
TIL Slutt BLE DNA ekstrahert fra frossen hjerne behandlet AV PCR for påvisning av bovin herpesvirus 1 og 5 (BHV-1 og -5), som tidligere beskrevet (Ashbaugh et al., 1997).
Bakteriologi
Ferske prøver av cerebrum og hjernestamme ble rutinemessig behandlet for aerobe bakteriekulturer i blod Og MacConkey agarer, og selektiv kultur For Listeria monocytogenes (Al-Zoreky og Sandine, 1990).
Resultater Og Diskusjon
de kliniske tegnene og epidemiologiske funnene i tilfellet beskrevet her, om enn ikke-spesifikt, var lik de som er beskrevet i andre tilfeller av bovin astrovirus-assosiert encefalitt, som vanligvis beskrives som sporadisk (Selimovic-Hamza et al ., 2016), med en rekke nevrologiske underskudd (Deiss et al., 2017), med en varighet av kliniske tegn som vanligvis varierer fra 1 dag til 3 uker (Schlottau et al ., 2016; Deiss et al., 2017; Stig et al., 2017; Hirashima et al., 2018).
Makroskopisk undersøkelse av hjernen, C1-segmentet i ryggmargen og andre vev i hodet avslørte ikke signifikante brutto anatomiske lesjoner. Histologisk var det moderat til alvorlig, lymfocytisk, histiocytisk og plasmacytisk meningoencefalomyelitt som påvirket telencephalon (inkludert hjernehalvdelen og hippocampus), hjernestammen og det eneste undersøkte segmentet av ryggmargen. Lesjoner ble overveiende fordelt i det grå stoffet og begrensende områder av hvitt materiale. I berørte områder var det perivaskulær cuffing og lymfoplasmacytisk og histiocytisk betennelse og nevronnekrose / nevronofagi med gliose i tilstøtende neuropil. Det var satellitose av berørte, nekrotiske nevroner(Figur 1A-D–. Lesjonene var mye mindre hyppige og alvorlige i cerebellar parenchyma, selv om det var multifokal moderat cerebellar leptomeningitt. Ingen intralesional bakterier ble funnet Med H& E og Gram flekker. Ingen signifikante histologiske endringer ble funnet i de andre undersøkte vevene.

Figur 1. Histologiske lesjoner i hjernestammen (A, B) og hjernebarken (C, D)og påvisning Av BoAstV RNA i hjernebarken (E,G). Bilder (Ad) er deler av hjernen farget Med H& E; bilder E Og G er deler av hjernebarken demonstrere hybridisering ved hjelp av kromogen ISH Ved Hjelp Av BoAstV spesifikk sonde, counterstained med hematoxylin; bilde F Er en seriell del av cerebral cortex dapb probe (negativ kontroll), counterstained med hematoxylin. (A) et perivenulært rom blir markert utvidet av inflammatoriske celler (for det meste lymfocytter og histiocytter) som også infiltrerer den tilstøtende nevropilen. (B) nevronet i midten har hypereosinofil perikaryon og karyorrhexis (nekrose) og nevronlegemet er omgitt av økt antall gliale (satellitose) og inflammatoriske celler. (C) leptomeninge infiltreres av lymfocytter og histiocytter. (D) en region av cerebral cortex med flere hypereosinofile (nekrotiske) nevroner og et stort fartøy med perivaskulære lymfocytiske mansjetter. (E) i en seriell del av hjernebarken er den rike Intracytoplasmatiske BoAstV RNA-merkingen avbildet ved sterk, granulær rød kromogenavsetning i neuronal cytoplasma av soma og neuronal extensions (E,G), som ikke er tilstede ved hybridisering ved bruk av negativ kontrollprobe (F).
en nevroinvasiv virusinfeksjon ble mistenkt ved histologisk undersøkelse av sentralnervesystemet. Storfe med encefalitt er bekymret fordi mange drøvtyggere nevropatogener er zoonotiske (Cantile og Youssef, 2016); dermed bør en diagnose av encefalitt føre til omfattende laboratorietesting for å skjerme for smittsomme stoffer når det er mulig. I det tilfellet som er beskrevet her, IHCs FOR WNV, rabiesvirus Og Chlamydia Spp., OG PCR FOR BHV-1 og -5 var alle negative, og ingen patogene bakterier ble dyrket fra hjernevev. Fordi steer var < 2 år gammel og ingen spongiform endringer ble observert i hjernestammen, ble dyret ikke testet for bovin spongiform encefalopati (BSE), som er en eksotisk sykdom hos voksne storfe som aldri har blitt rapportert i Uruguay. VIDERE ER BSE ikke inflammatorisk (Cantile og Youssef, 2016).
in situ hybridisering ble utført ved hjelp av en sonde generert Fra BoAstV-NeuroS1, og det var probe hybridisering rikelig innenfor, og begrenset til cytoplasma av nevroner i hjernehalvdelen og hippocampus(Figur 1E-G–. I disse områdene probe hybridisering colocalized med nekrotiske nevroner og regioner av gliosis, uten probe hybridisering påvises i gliaceller, eller inflammatoriske celler i perivaskulær mansjetter. Ingen viral nukleinsyre ble påvist AV ISH i cerebellum, som bare hadde minimal inflammatoriske lesjoner i parenchyma, men moderat leptomeningitt eller hjernestammen, inkludert seksjoner med alvorlig betennelse. Dette betyr at topografisk påvisning av viral distribusjon VED ISH var mer begrenset enn encefalitt i de undersøkte seksjonene, som noen ganger er beskrevet i tilfeller Av BoAstV-CH13 / NeuroS1-assosiert encefalitt hos storfe (Selimovic-Hamza et al ., 2017a, b). En årsak til denne sporadiske mangelen på viral RNA-deteksjon i lesioned områder av hjernen kan være deteksjonsgrensen FOR ISH, eller klaring av viruset i betente områder av hjernen ved dødstidspunktet, som tidligere antydet (Selimovic-Hamza et al .( 2017b). Som forventet ble ingen sondehybridisering detektert AV ISH i hjernevævet brukt som negativ kontroll.
Astrovirus ble oppdaget I hjernen AV RT-PCR. Nesten fullstendig genomsekvensanalyse avslørte En Mamastrovirusstamme i CH13 / NeuroS1-kladen, vi kalt BoAstV-Neuro-Uy, sekvensen ble deponert I GenBank under tiltredelsesnummer MK386569. Den fylogenetiske analysen viste nærhet med andre neuroinvasive astrovirus i Virginia / Human-Mink-Sau (VA/HMO) klade (Figur 2), som inneholder mest kjente neuroinvasive astrovirus (Hirashima et al., 2018; Reuter et al., 2018). Den nesten komplette sekvensen Av BoAstV-Neuro-Uy er 6427 bp i lengde og har en sekvensidentitet på 94% Med kagoshimasr28-462 stamme. BoAstV-Neuro-Uy har lignende egenskaper som andre stammer AV avstamning CH13/NeuroS1: en 5 ‘ UTR region av 51 nt, ORF1a (protease) av 861 aminosyrer (aa), ORF1b av 523 aa (RNA-avhengig rna polymerase), OG ORF2 av 758 aa (kapsidprotein). Dessverre kunne 3 ‘ UTR ikke sekvenseres, men en poly(A) hale antas å være tilstede fordi oligo (dT)18 ble brukt til å skaffe cDNA. I tillegg er den heptameriske aaaaaac-sekvensen, et ribosomalt rammeskiftesignal, tilstede. P-avstander VED aminosyrenivået TIL ORF2 bekreftet tildelingen av denne stammen TIL CH13 / NeuroS1-kladen. P-avstander < 0.35 Mellom BoAstV-Neuro-Uy og andre medlemmer av denne kladen (Tabell 1) vil støtte en klassifisering av disse virusstammene innen en samme art; Mamastrovirus-13 har nylig blitt foreslått av andre forfattere (Donato Og Vijaykrishna, 2017; Hirashima et al., 2018), selv om bestemt art tildeling AV ICTV venter. Sonden som ble brukt TIL ISH, generert Fra BoAstV-NeuroS1, hadde 92,7% sekvensidentitet Med BoAstV-Neuro-Uy.
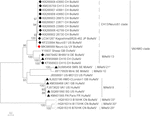
Figur 2. Fylogenetisk analyse av full lengde nukleotidsekvenser ved maksimal sannsynlighet metode. BoAstV-Neuro-Uy er merket med en rød rhombus. Andre sekvenser av bovin neuroinvasive astrovirus er merket med svart rhombi, sekvenser fra ikke-bovin neuroinvasive astrovirus er merket med svarte trekanter, og sekvenser fra enterisk bovin astrovirus er merket med hvite sirkler. Mamastrovirus arter er vist; stjerner indikerer arter som ennå ikke er anerkjent AV ICTV. Stammer INNENFOR CH13 / NeuroS1 klade ennå ikke tildelt en art AV ICTV er identifisert med en stiplet linje. Bootstrap verdier er angitt i noder. CH, Sveits; Jp, Japan; US, Amerikas Forente Stater; UY, Uruguay; GB, Det Forente Kongerike Storbritannia og NORD-IRLAND; DE, Tyskland; SE, Sverige; FR, Frankrike; Cn, Kina. BoAstV, bovint astrovirus; OvAstV, sau astrovirus; PoAstV, svin astrovirus; HuAstV, humant astrovirus; MiAstV, mink astrovirus.
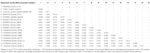
Tabell 1. Estimater av evolusjonære divergens på aminosyre nivå av hele ORF2 regionen mellom sekvenser av storfe Og sau Mamastrovirus-13 og stammer INNENFOR CH13 / NeuroS1 klade ennå ikke tildelt en art Av Den Internasjonale Komiteen For Taksonomi Av Virus (ICTV).
Studier basert på nevropatologiske undersøkelser og astrovirusnukleinsyre og proteindeteksjon har konkludert med at det er en sannsynlig årsakssammenheng mellom astrovirusinfeksjon og nevrologisk sykdom og lesjoner hos storfe (Selimovic-Hamza et al., 2017a; Reuter et al., 2018). Så vidt vi vet har astrovirus-assosiert encefalitt ikke blitt reprodusert eksperimentelt ennå. Dette ville kreve isolering av neuroinvasive astrovirus fra kliniske tilfeller, som ikke ble forsøkt i vårt tilfelle.
Kilden Til BoAstV-Neuro-Uy i dette tilfellet kunne ikke bestemmes. Derimot, reservoar storfe og dyreliv bør vurderes, som storfe ble oppdratt under omfattende utendørs forhold. Det berørte dyret var kjøpt og flyttet til gården i februar 2018 sammen med andre 9 styrer. Dessverre nektet eieren ytterligere prøvetaking og testing av andre dyr i eiendommen, og en mer detaljert epidemiologisk undersøkelse. Ingen av de andre dyrene i gruppen hadde utviklet nevrologisk sykdom fra August 2018, siste gang veterinæren ble kontaktet. En sesongmessighet fra begynnelsen av vinteren til slutten av våren har blitt foreslått for tilfeller av astrovirus-assosiert encefalitt I Sveits (Selimovic-Hamza et al., 2016). Interessant, saken beskrevet her skjedde i juni, tilsvarende høst-vinter overgangsperioden På Den Sørlige halvkule.
mens neurotropiske astrovirus har blitt identifisert I Nord-Amerika (Li et al., 2013; Spitsbergen et al., 2017), Europa (Bouzalas et al., 2014) Og Asia (Hirashima et al., 2018), har deres tilstedeværelse aldri blitt rapportert På Den Sørlige halvkule, så denne kommunikasjonen utvider den geografiske fordelingen av astrovirus-assosiert encefalitt. For å vurdere om virusstammen oppdaget I Uruguay kan ha sin opprinnelse I Europa, Nord-Amerika eller Asia, estimerte vi evolusjonsraten og utførte en fylogeografisk analyse ved hjelp av neuroinvasive BoAstV-sekvenser tilgjengelig I GenBank. Den evolusjonære hastighet estimert ved hjelp av komplett koding regionen var 4.27 × 10-4 (95% høyeste sannsynlighetstetthet-HPD-, 2.19–6.46 × 10-4) nukleotidsubstitusjoner/sted/år, som forventes for ET RNA-virus (Jenkins et al., 2002), men lavere enn estimert for enteriske humane astrovirus (Babkin et al., 2012, 2014). Den ORF1ab regionen viste en lignende evolusjonære pris (4.20 × 10-4, 95% HPD 1.66–6.46 × 10-4 innbytter/site/år) som fullstendig koding for regionen, mens ORF1a (2.92 × 10-4, 95% HPD 1.19 × 10-6-6.46 × 10-4 innbytter/site/år) og ORF2 (2.86 × 10-4, 95% HPD 4.13 × 10-6-5.79 × 10-4 substitusjoner/nettsted/år) viste en litt raskere evolusjonær rate, og Den partielle polymerase genomiske regionen (ORF1b) viste en litt langsommere evolusjonær rate (5.39 × 10-4, 95% HPD 6.41 × 10-7-1.10 × 10-3 substitusjoner/nettsted / år).
som bestemt av den fylogeografiske analysen med den komplette kodingsområdet, og vist I MCTT (Figur 3), er det to underlinjer (CH13 og NeuroS1) basert på referansestammer, som har en felles forfedre. Den siste felles stamfar til disse sub-linjene (lineage CH13/NeuroS1) oppsto i Europa omtrent i 1885 (95% HPD, 1794-1940). På begynnelsen av 1900-tallet divergerte DE to underlinjene, CH13-underlinjen ble sirkulerende i Europa, mens NeuroS1-underlinjen spredte Seg Til Amerika og Asia. Det mest sannsynlige scenariet er At NeuroS1-underlinjen ble introdusert I Uruguay fra Europa rundt år 1921 (95% HPD, 1849-1967), antagelig gjennom husdyrhandel, deretter spredt Til Nord-Amerika, og senere Til Japan (Figur 3). På grunn Av begrensningen i antall sekvenser tilgjengelig I GenBank, som kunne ha partisk analysen, ble resultatene oppnådd ved bruk av den komplette kodingsregionen sammenlignet med de som ble oppnådd med andre genomiske regioner (ORF1ab, ORF2, ORF1a og ORF1b) tilgjengelig for et større antall stammer (dvs.Kanadiske stammer). I alle analysene er det mest sannsynlige scenariet at innføringen av viruset til Uruguay skjedde Fra Europa (Supplerende Tall S1A–D). I tillegg var estimert dato for denne introduksjonen, oppnådd Med ORF1ab og partiell polymerase genomisk region (ORF1b) (Supplerende Tall S1A,D), lik den som ble oppnådd med den komplette kodingsregionen, mens estimert dato for introduksjon oppnådd MED ORF2 Og ORF1a var tidligere, men med bredere 95% HPD-intervall (Supplerende Tall S1B,C). En innføring av sub-avstamning NeuroS1 direkte Til Canada Fra Europa, med påfølgende spredning Til Usa Og Japan, er også plausibel, s vist I Supplerende Figur S1D.

Figur 3. Maksimal clade troverdighet treet oppnadd ved analyse av full lengde koding regionen. Fargen på grenene representerer det mest sannsynlige landet der forfedrene sirkulerte, bakre sannsynlighetsverdier er vist i grenene og tallene i hver node representerer opprinnelsesårene for hver klade med 95% HPD-intervallet. Underlinjer er angitt med etiketter.
Ytterligere undersøkelser er nødvendig for å vurdere geografisk fordeling, patogene mekanismer (spesielt mekanismer for overføring og oppføring), molekylær epidemiologi og potensiell interspecies overføring av neuroinvasive astrovirus.
Datatilgjengelighet
datasettene som genereres for denne studien, finnes I GenBank, MK386569.
Forfatterbidrag
FG, RDC og MC bidro med oppfatningen av studien. FG og RDC utførte patologisk undersøkelse og prøvetaking. PP utførte in situ hybridisering. FU utførte immunhistokjemi. LM, RC og MC utførte molekylær virologi testing. MC utførte sekvens-og fylogeografiske analyser og tilhørende tall. FG og PP fikk de histologiske bildene. MF utførte bakteriekulturer. FG og MC skrev det første utkastet til manuskriptet. RDC, PP, FU, LM, MF, og RC skrev deler av manuskriptet. Alle forfattere bidro til manuskriptrevisjon, leste og godkjente den innsendte versjonen.
Finansiering
dette arbeidet ble finansiert Av Tilskudd PL-015 N-15156 FRA Inia og 158 fra «Programa De Iniciaci@n a la Investigaci Chennayn 2017 «fra» Comisi@n Sectorial De Investigaci Chennayn Cientí » (CSIC). MC og RDC anerkjenner støtte fra HENHOLDSVIS «National Agency for Research and Innovation» (ANII) og inia gjennom Ph. d. stipendier. FG anerkjenner støtte FRA ANII gjennom mobility grant MOV_CA_2018_1_150021.
Interessekonflikt
forfatterne erklærer at forskningen ble utført i fravær av kommersielle eller økonomiske forhold som kan tolkes som en potensiell interessekonflikt.
anmelderen TS erklærte et tidligere medforfatterskap med EN av FORFATTERNE PP til håndteringsredaktøren.
Takk
forfatterne takker Yisell Perdomo Og Cecilia Monesiglio FRA INIA, Og Karen Sverlow Og Juliann Beingesser fra CAHFS for teknisk assistanse.
Supplerende Materiale
Supplerende Materiale for denne artikkelen kan bli funnet online på: https://www.frontiersin.org/articles/10.3389/fmicb.2019.01240/full#supplementary-material
FIGUR S1 / Maksimal clade troverdighet trær (MCCTs) oppnådd ved analyse av Full lengde ORF1ab (A), FULL lengde ORF2 (B), Full lengde ORF1a (C) og delvis ORF1b (D). Fargen på grenene representerer det mest sannsynlige landet der forfedrene sirkulerte, bakre sannsynlighetsverdier er vist i grenene, og tallene i hver node representerer opprinnelsesårene for hver klade med 95% HPD-intervallet. Underlinjer er angitt med etiketter.
Fotnoter
- ^http://iqtree.cibiv.univie.ac.at
Al-Zoreky, N. Og Sandine, W. E. (1990). Meget selektivt medium for isolering Av Listeria monocytogenes fra mat. Appl. Miljø. Mikrobiol. 56, 3154–3157.
PubMed Abstract | Google Scholar
Ashbaugh, S. E., Thompson, K. E., Belknap, E. B., Schultheiss, P. C., Chowdhury, S. og Collins, J. K. (1997). Spesifikk påvisning av shedding og latens av bovint herpesvirus 1 og 5 ved bruk av en nestet polymerasekjedereaksjon. J. Vet. Diagn. Investere. 9, 387–394. doi: 10.1177/104063879700900408
PubMed Abstract | CrossRef Full Text | Google Scholar
Babkin, I. V., Tikunov, A. Y., Sedelnikova, D. A., Zhirakovskaia, E. V., Og Tikunova, N. v. (2014). Rekombinasjonsanalyse basert På HAstV – 2 og HastV-4 komplette genomer. Infisere. Genet. Evol. 22, 94–102. doi: 10.1016 / j.meegid.2014.01.010
PubMed Abstract | CrossRef Full Text | Google Scholar
Babkin, I. V., Tikunov, A. Y., Zhirakovskaia, E. V., Netesov, S. V., Og Tikunova, N. v. (2012). Høy evolusjonær hastighet av menneskelig astrovirus. Infisere. Genet. Evol. 12, 435–442. doi: 10.1016 / j.meegid.2012.01.019
PubMed Abstract | CrossRef Full Text | Google Scholar
Blomstr@m, A. L., Bredé, F., Hammer, A. S., Belá, S., Og Berg, M. (2010). Påvisning av et nytt astrovirus i hjernevev av mink som lider av risting av mink syndrom ved bruk av viral metagenomikk. J. Clin. Mikrobiol. 48, 4392–4396. doi: 10.1128 / JCM.01040-10
PubMed Abstract | CrossRef Full Text | Google Scholar
Boros, Hryvnias, Albert, M., Pankovics, P., Bíó, H., Pesavento, P. A., Phan, T. G., et al. (2017). Utbrudd av neuroinvasiv astrovirus assosiert med encefalomyelitt, svakhet og lammelse blant avvente griser, Ungarn. Emerg. Infisere. Dis. 23, 1982–1993. doi: 10.3201 / eid2312. 170804
PubMed Abstrakt | CrossRef Fulltekst | Google Scholar
A. V., Koch, M. C., W., W., Werder, S., Jakupovic, D., Bruggmann, R., et al. (2017). Indikasjon av kryss-arter overføring av astrovirus assosiert med encefalitt hos sau og storfe. Emerg. Infisere. Dis. 23, 1604–1608. doi: 10.3201 / eid2309.170168
PubMed Abstrakt / CrossRef Fulltekst / Google Scholar
Bouzalas, I. G., W ④rich, D., Selimovic-Hamza, S., Drö, C., Bruggmann, R. og Seuberlich, T. (2016). Fullgenombasert molekylær karakterisering av encefalittassosiert bovint astrovirus. Infisere. Genet. Evol. 44, 162-168. doi: 10.1016 / J. meegid.06/052/2016
PubMed Abstrakt / CrossRef Fulltekst / Google Scholar
Bouzalas, I. G., Wü, D., Walland, J., Drö, C., Zurbriggen, A., Vandevelde, M., Et al. (2014). Neurotropisk astrovirus hos storfe med ikke-suppurativ encefalitt I Europa. J. Clin. Mikrobiol. 52, 3318–3324. doi: 10.1128 / JCM.01195-14
PubMed Abstract | CrossRef Full Text | Google Scholar
Cantile, C., Og Youssef, S. (2016). «Nervesystem», i Jubb, Kennedy, Og Palmers Patologi Av Husdyr, 6. Edn, ed. M. G. Maxie (Saint Louis, MO: Elsevier), 205-406.
Google Scholar
Deiss, R., Selimovic-Hamza, S., Seuberlich, T., Og Meylan, M. (2017). Nevrologiske kliniske tegn hos storfe med astrovirus-assosiert encefalitt. J. Vet. Praktikant. Med. 31, 1209–1214. doi: 10.1111 / jvim.14728
PubMed Abstrakt | CrossRef Fulltekst | Google Scholar
Donato, C., Og Vijaykrishna, D. (2017). Det brede vertsområdet og genetisk mangfold av pattedyr og aviær astrovirus. Virus 9: E102. doi: 10.3390 / v9050102
PubMed Abstrakt | CrossRef Fulltekst | Google Scholar
Drummond, A. J., Suchard, Ma, Xie, D., Og Rambaut, A. (2012). Bayesiansk fylogenetikk Med Skjønnhet og DYRET 1.7. Mol. Biol. Evol. 29, 1969–1973. doi: 10.1093 / molbev / mss075
PubMed Abstrakt / CrossRef Fulltekst / Google Scholar
J., Anderson, M., Miller, M., Rowe, J., Sverlow, K., Vasquez, M., et al. (2016). Chlamydia pecorum: foster-og plasentalskader i sporadisk kaprinabort. J. Vet. Diagn. Investere. 28, 184–189. doi: 10.1177/1040638715625729
PubMed Abstrakt / Fulltekst / Google Scholar
Hirashima, Y., Okada, D., Shibata, S., Yoshida, S., Fujisono, S., Omatsu, T., et al. (2018). Hele genomanalyse av et nytt neurotropisk bovint astrovirus oppdaget i En Japansk svart styre med ikke-suppurativ encefalomyelitt I Japan. Arch. Virol. 163, 2805–2810. doi: 10.1007 / s00705-018-3898-3
PubMed Abstrakt / Fulltekst / Google Scholar
Den Internasjonale Komiteen For Taksonomi Av Virus (2018). Master Species List (MSL32), Versjon 1. Tilgjengelig på: https://talk.ictvonline.org/files/master-species-lists/m/msl/7185 (tilgjengelig Mars, 2019).
Jenkins, G. M., Rambaut, A., Pybus, O. G., Og Holmes, E. C. (2002). Priser på molekylær evolusjon I RNA-virus: en kvantitativ fylogenetisk analyse. J. Mol. Evol. 54, 156–165. doi: 10.1007 / s00239-001-0064-3
PubMed Abstrakt / Fulltekst / Google Scholar
Kumar, S., Stecher, G., Og Tamura, K. (2016). MEGA7: molekylær evolusjonær genetikk analyse versjon 7.0 for større datasett. Mol. Biol. Evol. 33, 1870–1874. doi: 10.1093 / molbev / msw054
PubMed Abstrakt / CrossRef Fulltekst / Google Scholar
Li, L., Diab, S., McGraw, S., Barr, B., Traslavina, R., Higgins, R., et al. (2013). Divergent astrovirus assosiert med nevrologisk sykdom hos storfe. Emerg. Infisere. Dis. 19, 1385–1392. doi: 10.3201 / eid1909. 130682
PubMed Abstrakt | CrossRef Fulltekst | Google Scholar
Lole, K. S., Bollinger, R. C., Paranjape, R. S., Gadkari, D., Kulkarni, S. S., Novak, N. G., Et al. (1999). Full lengde humant immunsviktvirus type 1 genomer fra subtype C-infiserte serokonvertere I India, med tegn på intersubtype rekombinasjon. J. Virol. 73, 152–160.
PubMed Abstract | Google Scholar
Naccache, S. N. I tillegg er det et stort antall av dem. (2015). Diagnose av neuroinvasiv astrovirusinfeksjon hos en immunkompromittert voksen med encefalitt ved upartisk neste generasjons sekvensering. Clin. Infisere. Dis. 60, 919–923. doi: 10.1093/cid / ciu912
PubMed Abstrakt / CrossRef Fulltekst / Google Scholar
Jørgen, C., Jørgen, M., Jørgen, M., Jørgen, m., jørgen, l., et al. (2011). Patologi og immunhistokjemiske funn Av West Nile virus infeksjon i psittaciformes. Veterinær. Pathol. 48, 975–984. doi: 10.1177/0300985810391112
PubMed Abstract | CrossRef Full Text | Google Scholar
Pfaff, F., Schlottau, K., Scholes, S., Courtenay, A., Hoffmann, B., Hö, D., Et al. (2017). En roman astrovirus assosiert med encefalitt og ganglioneuritt hos husdyr. Transgående. Emerg. Dis. 64, 677–682. doi: 10.1111 / tbed.12623
PubMed Abstrakt | CrossRef Fulltekst | Google Scholar
T. A., T. A., T. A., T. A., T. A., T. A., T. A., T. A., T. A., Et al. (2010). Astrovirus encefalitt hos gutt Med X-bundet agammaglobulinemi. Emerg. Infisere. Dis. 16, 918–925. doi: 10.3201 / eid1606. 091536
PubMed Abstrakt | Kryssref Full Tekst | Google Scholar
Rambaut, A., Lam, T. T., Max Carvalho, L., Og Pybus, O. G. (2016). Utforske den temporale strukturen av heterokrone sekvenser ved Hjelp Av TempEst (formely Path-O-Gen). Virus Evol. 2: vew007. doi: 10.1093/ve / vew007
PubMed Abstrakt | CrossRef Fulltekst | Google Scholar
Reuter, G., Pankovics, P., Og Boros, A. (2018). Ikke-suppurativ (aseptisk) meningoencefalomyelitt assosiert med neurovirulente astrovirusinfeksjoner hos mennesker og dyr. Clin. Mikrobiol. Åp 31, e00040-18. doi: 10.1128 / CMR.00040-18
PubMed Abstract | CrossRef Full Text | Google Scholar
Schlottau, K., Schulze, C., Bilk, S., Hanke, D., Hö, D., Beer, M., et al. (2016). Påvisning av et nytt bovint astrovirus i en ku med encefalitt. Transgående. Emerg. Dis. 63, 253–259. doi: 10.1111 / tbed.12493
PubMed Abstrakt | CrossRef Fulltekst | Google Scholar
Selimovic-Hamza, S., Boujon, C. L., Hilbe, M., Oevermann, A. ,Og Seuberlich, T. (2017a). Frekvens og patologisk fenotype av bovint astrovirus CH13 / NeuroS1 infeksjon i nevrologisk sykdom storfe: mot vurdering av årsakssammenheng. Virus 9: E12. doi: 10.3390 / v9010012
PubMed Abstrakt | CrossRef Fulltekst | Google Scholar
Selimovic-Hamza, S., Sanchez, S., Philibert, H., Clark, E. G., Og Seuberlich, T. (2017b). Bovin astrovirusinfeksjon i feedlot storfe med nevrologisk sykdom i vest-Canada. Kan. Veterinær. J. 58, 601-603.
Google Scholar
Selimovic-Hamza, S., Bouzalas, I. G., Vandevelde, M., Oevermann, A., Og Seuberlich, T. (2016). Påvisning av astrovirus i historiske tilfeller Av Europeisk sporadisk bovin encefalitt, Sveits 1958-1976. Front. Veterinær. Sci. 3:91. doi: 10.3389 / fvets.2016.00091
PubMed Abstract | CrossRef Full Text | Google Scholar
Seuberlich, T., W@thrich, D., Selimovic-Hamza, S., Drö Hryvller, C., Oevermann, A., Bruggmann, R., et al. (2016). Identifikasjon av et annet encefalittassosiert astrovirus hos storfe. Emerg. Mikrober Infiserer. 5: e5. doi: 10.1038 / emi.2016.5
PubMed Abstract | CrossRef Full Text | Google Scholar
Spinato, M. T., Vince, A., Cai, H., Og Ojkic, D. (2017). Identifikasjon av bovint astrovirus i tilfeller av bovin ikke-suppurativ encefalitt i øst-Canada. Kan. Veterinær. J. 58, 607-609.
Google Scholar
Stein, L. T., Rech, R. R., Harrison, L. Og Brown, C. C. (2010). Immunhistokjemisk studie av rabiesvirus i sentralnervesystemet hos husdyr og dyrearter. Veterinær. Pathol. 47, 630–633. doi: 10.1177/0300985810370013
PubMed Abstrakt / Fulltekst / Google Scholar
Trifinopoulos, J., Nguyen, L. T., von Haeseler, A., Og Minh, B. Q. (2016). W-IQ-TREE: en rask online fylogenetisk verktøy for maksimal sannsynlighet analyse. Nukleinsyrer Res. 44, W232-W235. doi: 10.1093 / nar / gkw256
PubMed Abstrakt | CrossRef Fulltekst | Google Scholar
Tse, H., Chan, W. M., Tsoi, H. W., Fan, r. Y., Lau, C. C., Lau, S. K., et al. (2011). Gjenoppdagelse og genomisk karakterisering av bovine astrovirus. J. Gen. Virol. 92 (Pt 8), 1888-1998. doi: 10.1099 / vir.0.030817-0
PubMed Abstrakt / CrossRef Full Tekst / Google Scholar