biologie pentru Majors II
rezultatele învățării
- înțelegeți sistemele de clasificare anterioare și emergente pentru viruși
deoarece majoritatea virusurilor au evoluat probabil din strămoși diferiți, metodele sistematice pe care oamenii de știință le-au folosit pentru a clasifica celulele procariote și eucariote nu sunt foarte utile. Dacă virușii reprezintă” rămășițe ” ale diferitelor organisme, atunci nici analiza genomică sau proteică nu este utilă. De ce?, Deoarece virușii nu au o secvență genomică comună pe care o împărtășesc cu toții. De exemplu, secvența ARNr 16S atât de utilă pentru construirea filogeniilor procariote nu este de nici un folos pentru o creatură fără ribozomi! Biologii au folosit mai multe sisteme de clasificare în trecut. Virușii au fost inițial grupați după morfologie comună. Mai târziu, grupurile de viruși au fost clasificate după tipul de acid nucleic pe care îl conțineau, ADN sau ARN și dacă acidul lor nucleic era monocatenar sau dublu catenar. Cu toate acestea, aceste metode de clasificare anterioare grupau virușii în mod diferit, deoarece se bazau pe seturi diferite de caractere ale virusului. Cea mai frecvent utilizată metodă de clasificare astăzi se numește schema de clasificare Baltimore și se bazează pe modul în care ARN mesager (ARNm) este generat în fiecare tip particular de virus.
sistemele anterioare de clasificare
virusurile conțin doar câteva elemente prin care pot fi clasificate: genomul viral, tipul de capsidă și structura învelișului pentru virusurile învelite. Toate aceste elemente au fost utilizate în trecut pentru clasificarea virală (Tabelul 1 și Figura 1). Genomii virali pot varia în funcție de tipul de material genetic (ADN sau ARN) și de organizarea acestuia (monocatenar sau dublu catenar, liniar sau circular și segmentat sau nesegmentat). În unele virusuri, proteinele suplimentare necesare pentru replicare sunt asociate direct cu genomul sau conținute în capsida virală.
| Tabelul 1. Clasificarea virusului după structura și nucleul genomului | |
|---|---|
| clasificări de bază | Exemple |
| ARN | virusul rabiei, retrovirusuri |
| ADN | herpesvirusuri, virusul variolei |
| monocatenar | virusul rabiei, retrovirusuri |
| dublu catenar | herpesvirusuri, virusul variolei |
| liniar | virusul rabiei, retrovirusuri, herpesvirusuri, virusul variolei |
| circulară | papilomavirusuri, mulți bacteriofagi |
| non-segmentat: genomul constă dintr-un singur segment de material genetic | virusuri Parainfluenza |
| segmentat: genomul este împărțit în mai multe segmente | virusuri gripale |
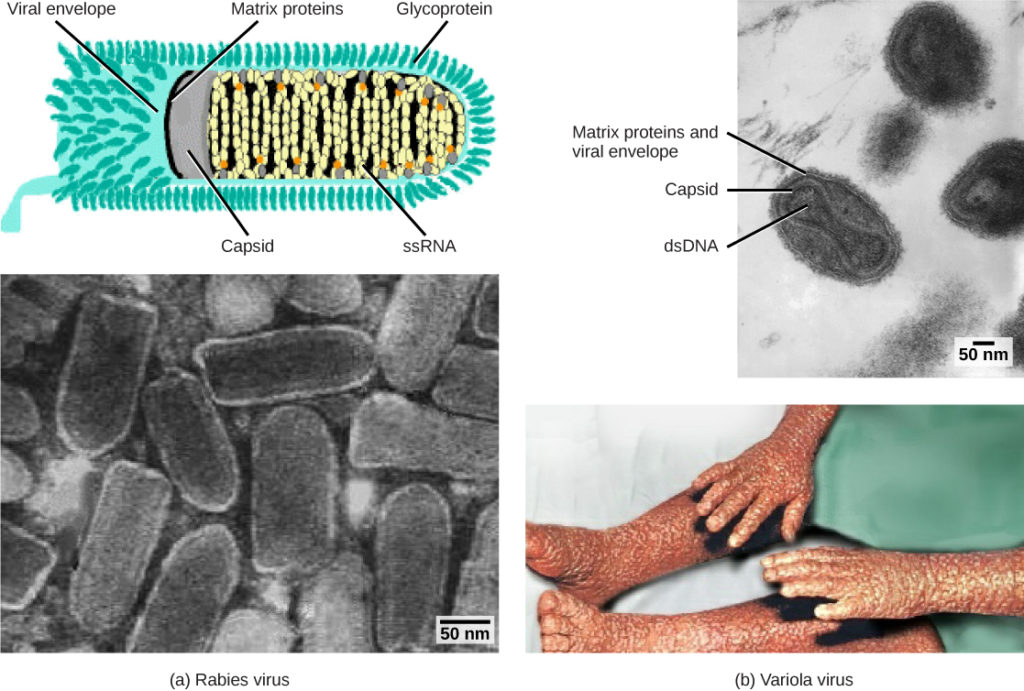
Figura 1. Virușii sunt clasificați pe baza materialului genetic de bază și a designului capsidei. (a) virusul rabiei are un nucleu ARN monocatenar (ssRNA) și o capsidă elicoidală învelită, în timp ce (B) virusul variolei, agentul cauzal al variolei, are un nucleu ADN dublu catenar (dsDNA) și o capsidă complexă. (credit „diagrama rabiei”: modificarea muncii de către CDC; „micrograful rabiei”: modificarea muncii de către Dr. Fred Murphy, CDC; credit „micrograful variolei mici”: modificarea muncii de către Dr. Fred Murphy, Sylvia Whitfield, CDC; credit „variola photo”: modificarea muncii de către CDC; date despre bara de scară de la Matt Russell)
virușii pot fi, de asemenea, clasificați prin proiectarea capsidelor lor (Tabelul 2 și Figura 2). Capsidele sunt clasificate ca icosaedrice goale, icosaedrice învelite, elicoidale învelite, elicoidale goale și complexe. Tipul de material genetic (ADN sau ARN) și structura acestuia (monocatenar sau dublu catenar, liniar sau circular și segmentat sau nesegmentat) sunt utilizate pentru clasificarea structurilor de bază ale virusului (Tabelul 2).
| Tabelul 2. Clasificarea virusului prin structura capsidei | |
|---|---|
| Clasificarea capsidei | Exemple |
| gol icosaedric | virusul hepatitei A, poliovirusuri |
| Icosaedric învelit | virusul Epstein-Barr, virusul herpes simplex, virusul rubeolei, virusul febrei galbene, HIV-1 |
| învelit elicoidal | virusurile gripale, virusul oreionului, virusul rujeolic, virusul rabiei |
| Naked elicoidal | virusul mozaicului tutunului |
| Complex cu multe proteine; unele au combinații de structuri capsidice icosaedrice și elicoidale | herpesvirusuri, virusul variolei, virusul hepatitei B, bacteriofagul T4 |

Figura 2. Micrografele electronice de transmisie ale diferitelor virusuri arată structurile lor. Capsida virusului poliomielitei (a) este icosaedrică goală; (b) capsida virusului Epstein-Barr este icosaedrică învelită; (c) capsida virusului oreionului este o helix învelită; (d) capsida virusului mozaicului tutunului este elicoidală goală; și (e) capsida herpesvirusului este complexă. (credit a: modificarea lucrărilor de Dr. Fred Murphy, Sylvia Whitfield; credit b: modificarea lucrărilor de Liza Gross; credit c: modificarea lucrărilor de Dr. F. A. Murphy, CDC; credit d: modificarea lucrărilor de USDA ARS; credit e: modificarea lucrării de Linda Stannard, Departamentul de Microbiologie medicală, Universitatea din Cape Town, Africa de Sud, NASA; date de scară de la Matt Russell)
Clasificarea Baltimore
cel mai frecvent utilizat sistem de clasificare a virusului a fost dezvoltat de biologul câștigător al Premiului Nobel David Baltimore la începutul anilor 1970. pe lângă diferențele de morfologie și genetică menționate mai sus, schema de clasificare Baltimore grupează virușii în funcție de modul în care ARNm este produs în timpul ciclului replicativ al virusului.
virusurile din grupa I conțin ADN dublu catenar (dsDNA) ca genom. ARNm-ul lor este produs prin transcriere în același mod ca și în cazul ADN-ului celular.
virusurile din grupa II au ADN monocatenar (ssDNA) ca genom. Ei își convertesc genomii monocatenari într-un intermediar dsDNA înainte de a putea apărea transcrierea în ARNm.
virusurile din grupul III utilizează dsRNA ca genom. Lanțurile se separă, iar una dintre ele este utilizată ca șablon pentru generarea ARNm folosind ARN polimeraza dependentă de ARN codificată de virus.
virusurile din grupul IV au ssRNA ca genom cu polaritate pozitivă. Polaritatea pozitivă înseamnă că ARN-ul genomic poate servi direct ca ARNm. Intermediarii dsRNA, numiți intermediari replicativi, se fac în procesul de copiere a ARN genomic. Din acești intermediari se formează mai multe fire de ARN de lungime completă de polaritate negativă (complementară ARN genomic cu catenă pozitivă), care pot servi apoi ca șabloane pentru producerea de ARN cu polaritate pozitivă, incluzând atât ARN genomic de lungime completă, cât și ARNm virale mai scurte.
virusurile din grupa V conțin genomi ssRNA cu polaritate negativă, ceea ce înseamnă că secvența lor este complementară ARNm. Ca și în cazul virusurilor din grupa IV, intermediarii dsRNA sunt utilizați pentru a face copii ale genomului și pentru a produce ARNm. În acest caz, genomul cu catenă negativă poate fi convertit direct în ARNm. În plus, firele ARN pozitive de lungime completă sunt realizate pentru a servi drept șabloane pentru producerea genomului cu catenă negativă.
virusurile din grupa VI au genomi diploizi (două copii) ssRNA care trebuie convertiți, folosind enzima revers transcriptază, în dsDNA; dsDNA este apoi transportat la nucleul celulei gazdă și introdus în genomul gazdă. Apoi, ARNm poate fi produs prin transcrierea ADN-ului viral care a fost integrat în genomul gazdă.
virusurile din grupa VII au genomi parțiali dsDNA și fac intermediari ssRNA care acționează ca ARNm, dar sunt, de asemenea, convertiți înapoi în genomi dsDNA prin transcriptază inversă, necesară pentru replicarea genomului. Caracteristicile fiecărui grup din clasificarea Baltimore sunt rezumate în tabelul 3 cu exemple ale fiecărui grup.
| Tabelul 3. Clasificarea Baltimore | |||
|---|---|---|---|
| Grupa | caracteristici | modul de producere a ARNm | exemplu |
| I | ADN dublu catenar | ARNm este transcris direct din șablonul ADN | Herpes simplex (herpesvirus) |
| II | ADN monocatenar | ADN-ul este transformat în formă dublu catenară înainte ca ARN-ul să fie transcris | parvovirus Canin (parvovirus) |
| III | ARN dublu catenar | ARNm este transcris din genomul ARN | gastroenterita copilăriei (rotavirus) |
| IV | ARN monocatenar (+) | genomul funcționează ca ARNm | răceală comună (pircornavirus) |
| V | ARN monocatenar (−) | ARNm este transcris din genomul ARN | rabie (rabdovirus) |
| VI | virusuri ARN monocatenare cu revers transcriptază | revers transcriptaza face ADN din genomul ARN; ADN-ul este apoi încorporat în genomul gazdă; ARNm este transcris din ADN-ul încorporat | virusul imunodeficienței umane (HIV) |
| VII | virusuri ADN dublu catenare cu revers transcriptază | genomul viral este ADN dublu catenar, dar ADN-ul viral este replicat printr – un ARN intermediar; ARN-ul poate servi direct ca ARNm sau ca șablon pentru a face ARNm | virusul hepatitei B (hepadnavirus) |