Biologi för Majors II
lärandemål
- förstå tidigare och framväxande klassificeringssystem för virus
eftersom de flesta virus troligen utvecklats från olika förfäder, är de systematiska metoder som forskare har använt för att klassificera prokaryota och eukaryota celler inte särskilt användbara. Om virus representerar” rester ” av olika organismer, är inte ens genomisk eller proteinanalys användbar. Varför?, Eftersom virus inte har någon gemensam genomisk sekvens som de alla delar. Till exempel är 16S rRNA-sekvensen så användbar för att konstruera prokaryotfylogenier inte till nytta för en varelse utan ribosomer! Biologer har tidigare använt flera klassificeringssystem. Virus grupperades ursprungligen efter delad morfologi. Senare klassificerades grupper av virus efter den typ av nukleinsyra de innehöll, DNA eller RNA, och om deras nukleinsyra var enkel – eller dubbelsträngad. Dessa tidigare klassificeringsmetoder grupperade emellertid virus på olika sätt, eftersom de baserades på olika uppsättningar av tecken i viruset. Den vanligaste klassificeringsmetoden idag kallas Baltimore classification scheme, och är baserad på hur messenger RNA (mRNA) genereras i varje viss typ av virus.
tidigare klassificeringssystem
virus innehåller endast några få element genom vilka de kan klassificeras: virusgenomet, typen av kapsid och kuvertstrukturen för de höljda virusen. Alla dessa element har tidigare använts för viral klassificering (Tabell 1 och figur 1). Virala genom kan variera i typen av genetiskt material (DNA eller RNA) och dess organisation (enkel – eller dubbelsträngad, linjär eller cirkulär och segmenterad eller icke-segmenterad). I vissa virus är ytterligare proteiner som behövs för replikering associerade direkt med genomet eller inneslutna i viruskapsiden.
| Tabell 1. Virusklassificering genom Genomstruktur och kärna | |
|---|---|
| kärn klassificeringar | exempel |
| RNA | rabiesvirus, Retrovirus |
| DNA | herpesvirus, smittkoppsvirus |
| enkelsträngat | rabiesvirus, Retrovirus |
| dubbelsträngat | herpesvirus, smittkoppsvirus |
| linjärt | rabiesvirus, Retrovirus, herpesvirus, smittkoppsvirus |
| cirkulär | papillomavirus, många bakteriofager |
| icke-segmenterad: genom består av ett enda segment av genetiskt material | parainfluensavirus |
| segmenterad: genomet är uppdelat i flera segment | influensavirus |
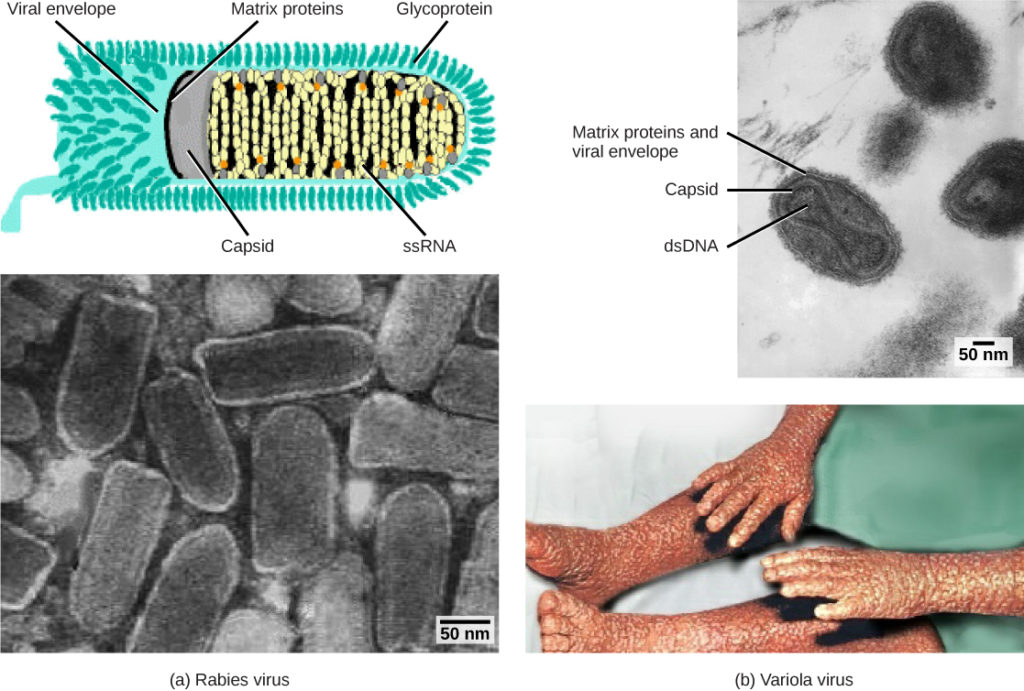
Figur 1. Virus klassificeras baserat på deras kärngenetiska material och kapsiddesign. (a) rabiesvirus har en enkelsträngad RNA-kärna (ssRNA) och en höljd spiralformad kapsid, medan (b) variolavirus, smittkoppens orsaksmedel, har en dubbelsträngad DNA-kärna (dsDNA) och en komplex kapsid. (kredit ”rabies diagram”: modifiering av arbete av CDC; ”rabies micrograph”: modifiering av arbete av Dr. Fred Murphy, CDC; kredit ”small pox micrograph”: modifiering av arbete av Dr. Fred Murphy, Sylvia Whitfield, CDC; kredit ”smittkoppor foto”: modifiering av arbete av CDC; scale-bar data från Matt Russell)
virus kan också klassificeras genom utformningen av deras capsids (Tabell 2 och figur 2). Capsids klassificeras som nakna icosahedral, omslagen icosahedral, omslagen spiralformad, naken spiralformad och komplex. Typen av genetiskt material (DNA eller RNA) och dess struktur (enkel – eller dubbelsträngad, linjär eller cirkulär och segmenterad eller icke-segmenterad) används för att klassificera viruskärnstrukturerna (Tabell 2).
| Tabell 2. Virusklassificering med Kapsidstruktur | |
|---|---|
| Capsid klassificering | exempel |
| naken icosahedral | hepatit A-virus, poliovirus |
| höljd ikosaedrisk | Epstein-Barr-virus, herpes simplexvirus, rubellavirus, gula febervirus, HIV-1 |
| omsluten spiralformad | influensavirus, påssjukevirus, mässlingvirus, rabiesvirus |
| naken spiralformad | tobaksmosaikvirus |
| komplex med många proteiner; vissa har kombinationer av ikosaedriska och spiralformade kapsidstrukturer | herpesvirus, smittkoppsvirus, hepatit B-virus, T4-bakteriofag |

Figur 2. Transmissionselektronmikrografer av olika virus visar sina strukturer. Kapsiden för (a) polioviruset är naken icosahedral; (b) Epstein-Barr-viruskapsiden är omslagen icosahedral; (c) påssjuka-viruskapsiden är en omslagen helix; (d) tobaksmosaikviruskapsiden är naken spiralformad; och (e) herpesviruskapsiden är komplex. (kredit a: modifiering av arbete av Dr. Fred Murphy, Sylvia Whitfield; kredit b: modifiering av arbete av Liza Gross; kredit c: modifiering av arbete av Dr. F. A. Murphy, CDC; kredit d: modifiering av arbete av USDA ARS; kredit e: ändring av arbete av Linda Stannard, Institutionen för medicinsk mikrobiologi, University of Cape Town, Sydafrika, NASA; scale-bar data från Matt Russell)
Baltimore Classification
det vanligaste systemet för virusklassificering utvecklades av Nobelprisvinnande biolog David Baltimore i början av 1970-talet. förutom skillnaderna i morfologi och genetik som nämns ovan grupperar Baltimore-klassificeringsschemat virus enligt hur mRNA produceras under virusets replikativa cykel.
grupp i-virus innehåller dubbelsträngat DNA (dsDNA) som deras genom. Deras mRNA produceras genom transkription på ungefär samma sätt som med cellulärt DNA.
grupp II-virus har enkelsträngat DNA (ssDNA) som deras genom. De omvandlar sina enkelsträngade genom till en dsDNA-mellanprodukt innan transkription till mRNA kan inträffa.
Grupp III-virus använder dsRNA som deras genom. Strängarna separeras, och en av dem används som en mall för generering av mRNA med användning av det RNA-beroende RNA-polymeras som kodas av viruset.
grupp IV-virus har ssRNA som sitt genom med en positiv polaritet. Positiv polaritet innebär att det genomiska RNA kan fungera direkt som mRNA. Intermediärer av dsRNA, kallade replikativa intermediärer, görs i processen att kopiera det genomiska RNA. Multipla RNA-strängar i full längd med negativ polaritet (gratis till det positiva strängade genomiska RNA) bildas från dessa mellanprodukter, som sedan kan fungera som mallar för produktion av RNA med positiv polaritet, inklusive både genomiskt RNA i full längd och kortare virala mRNA.
grupp V-virus innehåller ssRNA-genom med negativ polaritet, vilket innebär att deras sekvens är komplementär till mRNA. Som med grupp IV-virus används dsRNA-intermediärer för att göra kopior av genomet och producera mRNA. I detta fall kan det negativa strängade genomet omvandlas direkt till mRNA. Dessutom görs positiva RNA-strängar i full längd för att fungera som mallar för produktion av det negativa strängade genomet.
grupp vi-virus har diploida (två kopior) ssRNA-genom som måste omvandlas med hjälp av enzymet omvänd transkriptas till dsDNA; dsDNA transporteras sedan till kärnan i värdcellen och sätts in i värdgenomet. Sedan kan mRNA produceras genom transkription av viralt DNA som integrerades i värdgenomet.
grupp VII-virus har partiella dsDNA-genom och gör ssRNA-intermediärer som fungerar som mRNA, men omvandlas också tillbaka till dsDNA-genom genom omvänd transkriptas, nödvändigt för genomreplikation. Egenskaperna för varje grupp i Baltimore-klassificeringen sammanfattas i tabell 3 med exempel på varje grupp.
| tabell 3. Baltimore klassificering | |||
|---|---|---|---|
| grupp | egenskaper | läge för mRNA-produktion | exempel |
| i | dubbelsträngat DNA | mRNA transkriberas direkt från DNA-mallen | Herpes simplex (herpesvirus) |
| II | enkelsträngat DNA | DNA omvandlas till dubbelsträngad form innan RNA transkriberas | hundparvovirus (parvovirus) |
| III | dubbelsträngat RNA | mRNA transkriberas från RNA-genomet | barndoms gastroenterit (rotavirus) |
| IV | enkelsträngat RNA ( + ) | genom fungerar som mRNA | vanlig förkylning (pircornavirus) |
| V | enkelsträngat RNA ( – ) | mRNA transkriberas från RNA-genomet | Rabies (rhabdovirus) |
| vi | enkelsträngade RNA-virus med omvänt transkriptas | omvänt transkriptas gör DNA från RNA-genomet; DNA införlivas sedan i värdgenomet; mRNA transkriberas från det inkorporerade DNA | humant immunbristvirus (HIV) |
| VII | dubbelsträngade DNA-virus med omvänt transkriptas | virusgenomet är dubbelsträngat DNA, men viralt DNA replikeras genom en RNA-intermediär; RNA kan tjäna direkt som mRNA eller som en mall för att göra mRNA | hepatit B-virus( hepadnavirus) |