gränser i mikrobiologi
introduktion
Astroviridae-familjen innehåller icke-höljda, positiva, enkelsträngade RNA-virus inom två släkten, Mamastrovirus och avastrovirus, som infekterar däggdjur respektive fåglar. För närvarande erkänner International Committee on Taxonomy of Viruses (International Committee on Taxonomy of Viruses , 2018) 19 arter, nämligen Mamastrovirus-1 till -19, inom Mamastrovirus-släktet; det finns emellertid många stammar som väntar på klassificering, av vilka några preliminärt anses vara nya arter (Donato och Vijaykrishna, 2017).
sedan 2010 har flera astrovirus alltmer erkänts som neuroinvasiva hos olika däggdjursarter, inklusive människor (Quan et al., 2010; Naccache et al., 2015), mink (Blomstru., 2010), nötkreatur (Li et al., 2013), får (Pfaff et al., 2017), och grisar (Boros et al., 2017). Efter initialt erkännande av bovint astrovirusassocierad encefalit hos nötkreatur i USA (Li et al., 2013), en retrospektiv studie i fall av sporadisk bovin encefalit av obestämd etiologi från Schweiz avslöjade att detta neuroinvasiva astrovirus hade gått oupptäckt i årtionden (Selimovic-Hamza et al., 2016). Även om epidemiologin och överföringsvägarna för dessa astrovirus är okända, har överföring mellan arter föreslagits baserat på den höga identitetsnivån (>98%), delad mellan neuroinvasiva astrovirus från nötkreatur och får vid nukleotid-och aminosyranivåer (Boujon et al., 2017).
bovint astrovirus (BoAstVs), benämnt BoAstV-NeuroS1 (Li et al., 2013) och BoAstV-CH13 (Bouzalas et al., 2014), hittades ursprungligen i hjärnan hos nötkreatur med icke-suppurativ encefalit i USA respektive Schweiz. Trots den olika nomenklaturen representerar båda virusen samma genotypart (Bouzalas et al., 2016; Selimovic-Hamza et al., 2017a) som fortfarande väntar på officiell klassificering av ICTV. År 2015 identifierades en tidigare okänd BoAstV-stam, med namnet BoAstV-CH15, i hjärnan hos kor med encefalit i Schweiz. Fullständig genom fylogenetisk jämförelse avslöjade ett närmare förhållande mellan BoAstV-CH15 och ett får astrovirus (OvAstV) än med BoAstV-CH13 (Seuberlich et al., 2016). Coinfektion med BoAstV-CH13 och BoAstV-CH15 dokumenterades också i ett fall (Seuberlich et al., 2016). Samma år i Tyskland, Schlottau et al. (2016) rapporterade ett nytt astrovirus, nämligen BoAstV-BH89/14, i en ko med encefalit, som var närmast relaterad till OvAstV och BoAstV-CH15. Därefter identifierades BoAstV-CH13/NeuroS1 2017 i fall av bovin encefalit i östra och västra Kanada (Spinato et al., 2017; Selimovic-Hamza et al., 2017b). År 2018 identifierades en ny neuroinvasiv BoAstV nära besläktad med nordamerikanska och Europeiska BoAstV-NeuroS1/BoAstV-CH13 i en steer med icke-suppurativ encefalomyelit i Japan, och förekomsten av intra-genotypisk rekombination mellan de nordamerikanska och europeiska stammarna föreslogs (Hirashima et al., 2018).
medan fall av astrovirusassocierad encefalit har rapporterats i Nordamerika, Europa och Asien, har deras närvaro aldrig dokumenterats på södra halvklotet. Här beskriver vi ett fall av astrovirusassocierad encefalit hos nötkreatur i Uruguay, vilket breddar den geografiska fördelningen och genetiska mångfalden av neuroinvasiva astrovirus och ger fylogeografiska bevis som tyder på att detta virus infördes i Amerika från Europa och senare spred sig till Asien.
material och metoder
historia och signalering
i juni 2018 utvecklade en 22 månader gammal Holstein-styrning i en grupp på 37 stutar på en gård på 300 hektar i Colonia, Uruguay, progressiva neurologiska sjunger inklusive ovanligt beteende, mållös gång, cirkling, ataxi, repetitiva och okoordinerade tungrörelser och liggande. Besättningen betade på en årlig havre betesmark och kompletterades med majsensilage. En presumtiv klinisk diagnos av cerebral listerios av veterinärläkaren ledde till behandling med penicillin och streptomycin, men djuret dog spontant efter en klinisk kurs som varade 3 dagar.
patologisk undersökning, in situ hybridisering (ISH) och immunhistokemi (IHC)
styrhuvudet avlägsnades från slaktkroppen och skickades till INIAS Veterinärdiagnostiska laboratorium (Animal Health Platform) för diagnostisk upparbetning. Hälften av hjärnan, ett kort segment av proximal cervikal ryggmärg (C1), trigeminal ganglion och rot av trigeminusnerven, spottkörtel, retrofaryngeal lymfkörtel, orofarynx, matstrupe, tunga och skelettmuskel, var nedsänkning-fixerad i 10% neutral buffrad formalin för 48-72 h. vävnader behandlades rutinmässigt för histologi, inbäddad i paraffin, mikrotom-sektionerad vid 4-5 OC och färgad med hematoxylin och eosin (H&E) och Gramfläckar.
kromogen ISH utfördes manuellt på 5 micrym-sektioner av formalinfixerad, paraffininbäddad (FFPE) hjärnstam, cerebrum och cerebellum på Superfrost plus-bilder (Thermo Fisher Scientific, Pittsburgh, PA, USA) med hjälp av Rnascope 2.5 Red assay kit (Cat #322360, Advanced Cell Diagnostics, Hayward, CA, USA) och BoAstV-sonden Cat. #406921. Sonden består av 20zz-par som riktar sig mot regionen 5232-6180 av viruset (GenBank KF233994.1). Varje sektion av vävnad på 5 kg förbehandlades med värme och proteas före sondhybridisering i 2 timmar vid 40 kg C och bearbetades enligt tillverkarens rekommendationer. Negativa kontroller som användes för validering av signal inkluderade en orelaterad (GC-innehållsmatchad) sondkörning på seriella sektioner och sonderande vävnad från oinfekterade djur. Slides motfärgades med hematoxylin och monterades med EcoMount (Biocare Medical, Concord, CA, USA).
dessutom utfördes IHC i FFPE-sektioner av hjärnstam, cerebrum och cerebellum, som tidigare beskrivits, för identifiering av West Nile-virus (WNV, Flavivirus) (Palmieri et al., 2011), rabiesvirus (Lyssavirus) (Stein et al., 2010) och Chlamydia spp. (Giannitti et al., 2016) antigener.
molekylär virologi
Nukleinsyraextraktion åstadkoms från ett poolat prov av frusen (-20 kcal C) hjärna med användning av MagMAX Nukleinsyraisoleringssats (Thermo Fisher Scientific). För astrovirusdetektering utfördes omvänd transkription (RT) med RevertAid omvänd transkriptas Bisexuell (Thermo Fisher Scientific) och slumpmässiga hexamerprimrar (Qiagen). PCR utfördes från cDNA med användning av Mangomix Bisexuell (Biolin) och primers som förstärker ett 432-nukleotidfragment av astroviruspolymerasgenen (Tse et al., 2011). PCR-produkten visualiserades i 2% agarosgel, renades med användning av PureLink Bisexual Quick Gel Extraction och PCR Purification Combo Kit (Invitrogen) och sekvenserades vid Macrogen Inc. (Seoul, Sydkorea). För astrovirus hela genomförstärkning, Maxima h Minus omvänd transkriptas(Thermo Fisher Scientific) och oligo (dT)18 för obtention av cDNA, och Mangomix Bisexuell (Bioline) eller Ranger DNA-polymeras (Bioline) med primers beskrivna av Hirashima et al., 2018, användes. PCR-produkterna visualiserades i 1-2% agarosgel, renades och sekvenserades som nämnts ovan. Sekvensmontering utfördes med SeqMan (Lasergene 8, DNASTAR). Tjugosex kompletta genomsekvenser av neuroinvasiv astrovirus från nötkreatur, får, grisar, människor och minkoch enteriskt bovint astrovirus tillgängligt i GenBank laddades ner och justerades med Clustal W i MEGA 7-programvara (Kumar et al., 2016). W-IQ-TREE1 (Trifinopoulos et al., 2016) användes för att bestämma den bästa passformen för sekvensutveckling (SYM+i+G4) och att konstruera ett fylogenetiskt träd med maximal sannolikhet med de nästan fullständiga sekvenserna av BoAstV som detekterades i detta fall och de kompletta sekvenserna som hämtades från GenBank, med hjälp av bootstrap som statistisk metod för att bedöma clades robusthet. Likhetsplott utfördes med SimPlot-programvara (Lole et al., 1999). P-avstånd vid aminosyranivån i ORF2 uppskattades med MEGA 7-programvara (Kumar et al., 2016).
dessutom utfördes en bayesisk fylogeografisk analys med BEAST v1.8.4-paketet (Drummond et al., 2012), med användning av: den fullständiga kodningsregionen för BoAstV CH13 / NeuroS1-härstamning, ORF1ab (icke-strukturella gener), ORF2 (strukturell gen), ORF1a (proteas) och en partiell ORF1b (polymeras genomisk region, för vilken kanadensiska stammar var tillgängliga), med alla sekvenser tillgängliga i GenBank (förra anslutningen 18 April 2019), för att bestämma evolutionshastigheten, de gemensamma förfädernas åldrar/år och den mest sannolika vägen för viral cirkulation per land (Schweiz, Uruguay, USA, Kanada och Japan). Brist på rekombination i datauppsättningen bestämdes med användning av Rekombinationsdetekteringsprogram 4. Substitutionsmodellen som bäst passar varje inriktning bestämdes med hjälp av MEGA 7-programvara genom Bayesian information criterion (BIC) – värden, och den temporala strukturen för varje dataset utvärderades med hjälp av TempEst (Rambaut et al., 2016). Den lognormala avslappnade molekylära klockan med Bayesian Skyline-analys valdes av Bayes Factor bland de olika kombinationerna av molekylära klockor och koalescerande trädpriorer som användes. Detekteringslandet användes som drag. Markov-kedjan Monte Carlo längd var 100 miljoner generationer, vilket säkerställde konvergensen av analysen, utvärderad i Tracer v1.6.0, och den bakre sannolikheten användes för att utvärdera klader. Den maximala clade trovärdighet träd (MCCT) erhölls med hjälp av treeannotator programvara från BEAST och visualiseras i FigTree v1.4.3.
slutligen behandlades DNA extraherat från frusen hjärna med PCR för detektering av bovint herpesvirus 1 och 5 (BHV-1 och -5), som tidigare beskrivits (Ashbaugh et al., 1997).
bakteriologi
färska prover av hjärnan och hjärnstammen behandlades rutinmässigt för aeroba bakteriekulturer i blod-och MacConkey-agarer och selektiv kultur för Listeria monocytogenes (Al-Zoreky och Sandine, 1990).
resultat och diskussion
de kliniska tecknen och epidemiologiska fynd i det fall som beskrivs häri, om än icke-specifika, liknade de som beskrivs i andra fall av bovin astrovirusassocierad encefalit, som vanligtvis beskrivs som sporadisk (Selimovic-Hamza et al., 2016), med olika neurologiska underskott (Deiss et al., 2017), med en varaktighet av kliniska tecken som vanligtvis sträcker sig från 1 dag till 3 veckor (Schlottau et al., 2016; Deiss et al., 2017; Spinato et al., 2017; Hirashima et al., 2018).
makroskopisk undersökning av hjärnan, C1-segmentet i ryggmärgen och andra vävnader i huvudet avslöjade inte signifikanta grova anatomiska lesioner. Histologiskt var det måttlig till svår, lymfocytisk, histiocytisk och plasmacytisk meningoencefalomyelit som påverkade telencephalon (inklusive hjärnhalvan och hippocampus), hjärnstammen och det enda undersökta segmentet av ryggmärgen. Lesioner fördelades övervägande i gråmaterialet och begränsande områden av vit substans. I drabbade områden fanns perivaskulär manschett och lymfoplasmacytisk och histiocytisk inflammation och neuronal nekros/neuronofagi med glios i intilliggande neuropil. Det fanns satellitos av drabbade, nekrotiska neuroner (figurerna 1a–D). Lesionerna var mycket mindre frekventa och svåra i cerebellär parenchyma, även om det fanns multifokal måttlig cerebellär leptomeningit. Inga intralesionala bakterier hittades med h&E och Gram fläckar. Inga signifikanta histologiska förändringar hittades i de andra undersökta vävnaderna.

Figur 1. Histologiska lesioner i hjärnstammen (A,B) och hjärnbarken (C,D) och detektion av BoAstV RNA i hjärnbarken (E,G). Bilder (A-D) är delar av hjärnan färgade med H& E; bilder E och G är sektioner av hjärnbarken som visar hybridisering med kromogen ISH med hjälp av BoAstV-specifik sond, motfärgad med hematoxylin; bild F är en seriell sektion av hjärnbarken DapB-sond (negativ kontroll), motfärgad med hematoxylin. (A) ett perivenulärt utrymme expanderas markant av inflammatoriska celler (mestadels lymfocyter och histiocyter) som också infiltrerar den intilliggande neuropilen. (B) neuronen i mitten har hypereosinofil perikaryon och karyorrhexis (nekros) och den neuronala kroppen omges av ökat antal glial (satellitos) och inflammatoriska celler. (C) leptomeninge infiltreras av lymfocyter och histiocyter. (D) en region i hjärnbarken med flera hypereosinofila (nekrotiska) neuroner och ett stort kärl med perivaskulära lymfocytiska manschetter. (E) i en seriell sektion av hjärnbarken avbildas den rikliga intracytoplasmiska BoAstV-RNA-märkningen av stark, granulär röd kromogenavsättning i neuronal cytoplasma av soma och neuronala förlängningar (t.ex.), som inte är närvarande med hybridisering med hjälp av den negativa kontrollproben (F).
en neuroinvasiv virusinfektion misstänktes vid histologisk undersökning av centrala nervsystemet. Nötkreatur med encefalit är oroande eftersom många idisslare neuropatogener är zoonotiska (Cantile och Youssef, 2016); således bör en diagnos av encefalit leda till omfattande laboratorietester för att undersöka smittämnen när det är möjligt. I det fall som beskrivs häri, IHCs för WNV, rabiesvirus och Chlamydia spp., och PCR för BHV-1 och -5 var alla negativa, och inga patogena bakterier odlades från hjärnvävnad. Eftersom styret var < 2 år gammalt och inga spongiforma förändringar observerades i hjärnstammen testades inte djuret för bovin spongiform encefalopati (BSE), vilket är en exotisk sjukdom hos vuxna nötkreatur som aldrig har rapporterats i Uruguay. Dessutom är BSE inte inflammatorisk (Cantile och Youssef, 2016).
in situ hybridisering utfördes med användning av en sond genererad från BoAstV-NeuroS1, och det fanns sondhybridisering riklig inom och begränsad till cytoplasman hos neuroner i hjärnhalvan och hippocampus (figurerna 1e–G). I dessa områden samlokaliserades sondhybridiseringen med nekrotiska neuroner och regioner av glios, utan sondhybridisering detekterbar i gliacellerna eller inflammatoriska celler i perivaskulära manschetter. Ingen viral nukleinsyra detekterades av ISH i cerebellum, som endast hade minimala inflammatoriska lesioner i parenkymen men måttlig leptomeningit eller hjärnstammen, inklusive sektioner med svår inflammation. Detta innebär att topografiskt var detektion av viral distribution av ISH mer begränsad än encefalit i de undersökta sektionerna, vilket ibland har beskrivits i fall av BoAstV-CH13/NeuroS1-associerad encefalit hos nötkreatur (Selimovic-Hamza et al., 2017a, b). En orsak till denna tillfälliga brist på viral RNA-detektion i skadade områden i hjärnan kan vara detektionsgränsen för ISH eller clearance av viruset i inflammerade områden i hjärnan vid dödsfallet, som tidigare föreslagits (Selimovic-Hamza et al., 2017b). Som förväntat detekterades ingen sondhybridisering av ISH i hjärnvävnaden som användes som negativ kontroll.
Astrovirus detekterades i hjärnan med RT-PCR. Nästan fullständig genomsekvensanalys avslöjade en Mamastrovirusstam inom CH13 / NeuroS1-kladen, vi namngav BoAstV-Neuro-Uy, sekvensen deponerades i GenBank under anslutningsnummer MK386569. Den fylogenetiska analysen avslöjade närhet till andra neuroinvasiva astrovirus inom Virginia/Human-Mink-får (VA/HMO) klad (Figur 2), som innehåller mest kända neuroinvasiva astrovirus (Hirashima et al., 2018; Reuter et al., 2018). Den nästan fullständiga sekvensen av BoAstV-Neuro-Uy är 6427 bp i längd och har en sekvensidentitet på 94% med kagoshimasr28-462-stam. BoAstV-Neuro-Uy har liknande egenskaper som andra stammar av härstamning CH13 / NeuroS1: en 5 ’ utr-region av 51 nt, ORF1a (proteas) av 861 aminosyror (aa), ORF1b av 523 aa (RNA-beroende RNA-polymeras) och ORF2 av 758 aa (kapsidprotein). Tyvärr kunde 3 ’ UTR inte sekvenseras, men en poly(a) svans antas vara närvarande eftersom oligo(dT)18 användes för att erhålla cDNA. Dessutom är den heptameriska aaaaaac-sekvensen, en ribosomal frameshift-signal, närvarande. P-avstånd vid aminosyranivån i ORF2 bekräftade tilldelningen av denna stam till CH13/NeuroS1-kladen. P-avstånd < 0.35 mellan BoAstV-Neuro-Uy och andra medlemmar av denna klad (Tabell 1) skulle stödja en klassificering av dessa virusstammar inom samma art; Mamastrovirus-13 har nyligen föreslagits av andra författare (Donato och Vijaykrishna, 2017; Hirashima et al., 2018), även om bestämd arttilldelning av ICTV väntar. Sonden som användes för ISH, genererad från BoAstV-NeuroS1, hade 92.7% sekvensidentitet med BoAstV-Neuro-Uy.
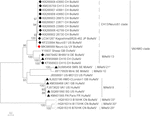
Figur 2. Fylogenetisk analys av nukleotidsekvenser i full längd med maximal sannolikhetsmetod. BoAstV-Neuro-Uy är märkt med en röd rhombus. Andra sekvenser av bovint neuroinvasiv astrovirus är märkta med svart romb, sekvenser från icke-bovint neuroinvasiv astrovirus är märkta med svarta trianglar, och sekvenser från enteriska bovint astrovirus är märkta med vita cirklar. Mamastrovirusarter visas; asterisker indikerar arter som ännu inte känns igen av ICTV. Stammar inom CH13 / NeuroS1-kladen som ännu inte tilldelats en art av ICTV identifieras med en streckad linje. Bootstrap-värden anges i noder. CH, Schweiz; JP, Japan; USA, Amerikas förenta stater; UY, Uruguay; GB, Förenade konungariket Storbritannien; De, Tyskland; se, Sverige; FR, Frankrike; CN, Kina. BoAstV, bovint astrovirus, OvAstV, får astrovirus, PoAstV, svin astrovirus, HuAstV, humant astrovirus, MiAstV, mink astrovirus.
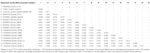
Tabell 1. Uppskattningar av evolutionär divergens vid aminosyranivån i hela ORF2-regionen mellan sekvenser av nötkreatur och får Mamastrovirus-13 och stammar inom ch13/NeuroS1-kladen som ännu inte tilldelats en art av International Committee on Taxonomy of Viruses (ICTV).
studier baserade på neuropatologiska undersökningar och astrovirusnukleinsyra och proteindetektion har dragit slutsatsen att det finns ett troligt orsakssamband mellan astrovirusinfektion och neurologisk sjukdom och lesioner hos nötkreatur (Selimovic-Hamza et al., 2017a; Reuter et al., 2018). Så vitt vi vet har astrovirusassocierad encefalit inte reproducerats experimentellt ännu. Detta skulle kräva isolering av neuroinvasiva astrovirus från kliniska fall, vilket inte försökte i vårt fall.
källan till BoAstV-Neuro-Uy i detta fall kunde inte bestämmas. Reservoarboskap och djurliv bör dock övervägas, eftersom nötkreaturen uppföddes under omfattande utomhusförhållanden. Det drabbade djuret hade köpts och flyttats till gården i februari 2018 tillsammans med andra 9 stutar. Tyvärr vägrade ägaren ytterligare provtagning och testning av andra djur i fastigheten och en mer detaljerad epidemiologisk undersökning. Inget av de andra djuren i gruppen hade utvecklat neurologisk sjukdom från och med augusti 2018, sista gången veterinärläkaren kontaktades. En säsongsmässighet från början av vintern till slutet av våren har föreslagits för fall av astrovirusassocierad encefalit i Schweiz (Selimovic-Hamza et al., 2016). Intressant är att det fall som beskrivs här inträffade i juni, vilket motsvarar höst-vinter övergångsperioden på södra halvklotet.
medan neurotropa astrovirus har identifierats i Nordamerika (Li et al., 2013; Spinato et al., 2017), Europa (Bouzalas et al., 2014) och Asien (Hirashima et al., 2018) har deras närvaro aldrig rapporterats på södra halvklotet, så denna kommunikation breddar den geografiska fördelningen av astrovirusassocierad encefalit. För att bedöma om virusstammen som upptäckts i Uruguay kan ha sitt ursprung i Europa, Nordamerika eller Asien uppskattade vi evolutionshastigheten och utförde en fylogeografisk analys med hjälp av neuroinvasiva BoAstV-sekvenser tillgängliga i GenBank. Den evolutionära hastigheten som uppskattades med hjälp av den fullständiga kodningsregionen var 4.27 10-4 (95% högsta sannolikhetstäthet-HPD -, 2,19-6,46 10-4 20-4) nukleotid substitutioner / plats / år, vilket förväntas för ett RNA-virus (Jenkins et al., 2002), men lägre än det som uppskattas för enteriska humana astrovirus (Babkin et al., 2012, 2014). Den ORF1ab regionen visade en liknande evolutionära ränta (4.20 × 10-4, 95% HPD 1.66–6.46 × 10-4 substitutioner/plats/år) som fullständig kodning regionen, medan den ORF1a (2.92 × 10-4, 95% HPD 1.19 × 10-6-6.46 × 10-4 substitutioner/plats/år) och ORF2 (2.86 × 10-4, 95% HPD 4.13 × 10-6-5.79 20-4 10-4 substitutioner/plats/år) visade en något snabbare evolutionär hastighet, och den partiella polymeras genomiska regionen (ORF1b) visade en något långsammare evolutionär hastighet(5,39 10-4, 95% HPD 6,41 10-7-1, 10 10-3 substitutioner/plats/år).
som bestäms av fylogeografisk analys med den fullständiga kodningsregionen och visas i MCTT (Figur 3) finns det två underlinjer (CH13 och NeuroS1) baserat på referensstammar, som har en gemensam förfader. Den senaste gemensamma förfadern till dessa underlinjer (lineage CH13/NeuroS1) uppstod i Europa ungefär 1885 (95% HPD, 1794-1940). I början av 1900-talet divergerade de två underlinjerna, ch13-underlinjen stannade kvar i Europa, medan NeuroS1-underlinjen spred sig till Amerika och Asien. Det mest troliga scenariot är att NeuroS1-underlinjen introducerades i Uruguay från Europa omkring år 1921 (95% HPD, 1849-1967), förmodligen genom boskapshandel, spred sig sedan till Nordamerika och senare till Japan (Figur 3). På grund av begränsningen i antalet sekvenser tillgängliga i GenBank, som kunde ha partisk analysen, jämfördes resultaten som erhölls med den fullständiga kodningsregionen med de som erhölls med andra genomiska regioner (ORF1ab, ORF2, ORF1a och ORF1b) tillgängliga för ett större antal stammar (dvs. kanadensiska stammar). I alla analyser är det mest troliga scenariot att införandet av viruset till Uruguay inträffade från Europa (kompletterande siffror S1A–D). Dessutom var det beräknade datumet för denna introduktion, erhållet med ORF1ab och partiell polymeras genomisk region (ORF1b) (kompletterande siffror S1A,D), liknande det som erhölls med den fullständiga kodningsregionen, medan det uppskattade datumet för introduktion erhållet med ORF2 och ORF1a var tidigare men med bredare 95% HPD-intervall (kompletterande siffror S1B,C). En introduktion av Sub-lineage NeuroS1 direkt till Kanada från Europa, med efterföljande spridning till USA och Japan, är också troligt, S visas i kompletterande figur S1D.

Figur 3. Maximal clade trovärdighet träd som erhållits genom analys av fullängds kodningsområdet. Färgen på grenarna representerar det mest troliga landet där förfäderna cirkulerade, bakre sannolikhetsvärden visas i grenarna och siffrorna i varje nod representerar ursprungsåren för varje klad med 95% HPD-intervallet. Underlinjer anges med etiketter.
ytterligare undersökningar behövs för att bedöma geografisk fördelning, patogena mekanismer (särskilt mekanismer för överföring och inträde), molekylär epidemiologi och potentiell överföring mellan arter av neuroinvasiva astrovirus.
datatillgänglighet
datauppsättningarna som genereras för denna studie finns i GenBank, MK386569.
Författarbidrag
FG, RDC och MC bidrog med uppfattningen av studien. FG och RDC utförde den patologiska undersökningen och provtagningen. PP utförde hybridiseringen in situ. FU utförde immunhistokemi. LM, RC och MC utförde molekylär virologi testning. MC utförde sekvensen och fylogeografiska analyser och tillhörande figurer. FG och PP fick de histologiska bilderna. MF utförde bakteriekulturerna. FG och MC skrev det första utkastet till manuskriptet. RDC, PP, FU, LM, MF och RC skrev delar av manuskriptet. Alla författare bidrog till manuskriptrevision, läste och godkände den inlämnade versionen.
finansiering
detta arbete har finansierats av bidrag PL-015 N-15156 från INIA och 158 från ”Programa de Iniciaci Ubicn a la investigaci Ubicn 2017 ”från” Comisi Ubicn Sectorial de investigaci Ubicn cient Ubicfica ” (CSIC). MC och RDC erkänner stöd från” National Agency for Research and Innovation ” (ANII) respektive Inia genom Ph.D. stipendier. FG erkänner stöd från ANII genom mobilitetsbidrag MOV_CA_2018_1_150021.
intressekonflikt uttalande
författarna förklarar att forskningen genomfördes i avsaknad av kommersiella eller finansiella relationer som kan tolkas som en potentiell intressekonflikt.
granskaren TS förklarade ett tidigare medförfattarskap med en av författarna PP till hanteringsredigeraren.
bekräftelser
författarna tackar Yisell Perdomo och Cecilia Monesiglio från INIA, och Karen Sverlow och Juliann Beingesser från CAHFS för teknisk hjälp.
tilläggsmaterial
Tilläggsmaterialet för denna artikel finns online på: https://www.frontiersin.org/articles/10.3389/fmicb.2019.01240/full#supplementary-material
figur S1 / maximal clade trovärdighet träd (MCCTs) som erhållits genom analys av full längd ORF1ab (a), full längd ORF2 (B), Full längd ORF1a (C), och partiell ORF1b (D). Färgen på grenarna representerar det mest troliga landet där förfäderna cirkulerade, bakre sannolikhetsvärden visas i grenarna och siffrorna i varje nod representerar ursprungsåren för varje klad med 95% HPD-intervallet. Underlinjer anges med etiketter.
fotnoter
- ^http://iqtree.cibiv.univie.ac.at
Al-Zoreky, N. och Sandine, W. E. (1990). Mycket selektivt medium för isolering av Listeria monocytogenes från mat. Appl. Miljö. Mikrobiol. 56, 3154–3157.
PubMed Abstrakt / Google Scholar
Ashbaugh, S. E., Thompson, K. E., Belknap, E. B., Schultheiss, P. C., Chowdhury, S. och Collins, J. K. (1997). Specifik detektion av utgjutning och latens av bovint herpesvirus 1 och 5 med användning av en kapslad polymeraskedjereaktion. J. Vet. Diagn. Investera. 9, 387–394. doi: 10.1177/104063879700900408
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Babkin, I. V., Tikunov, A. Y., Sedelnikova, D. A., Zhirakovskaia, E. V. och Tikunova, N. V. (2014). Rekombinationsanalys baserad på HAstV-2 och HastV-4 kompletta genom. Infektera. Genet. Evol. 22, 94–102. doi: 10.1016/j.meegid.2014.01.010
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Babkin, I. V., Tikunov, A. Y., Zhirakovskaia, E. V., Netesov, S. V. och Tikunova, N. V. (2012). Hög evolutionär hastighet av humant astrovirus. Infektera. Genet. Evol. 12, 435–442. doi: 10.1016/j.meegid.2012.01.019
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Blomstru, A. L., Widu, F., Hammer, A. S., Belu, S. och Berg, M. (2010). Detektion av ett nytt astrovirus i hjärnvävnad av mink som lider av skakande minksyndrom genom användning av viral metagenomik. J. Clin. Mikrobiol. 48, 4392–4396. doi: 10.1128/JCM.01040-10
PubMed Abstrakt / CrossRef Fulltext / Google Scholar
Boros, Baccarat, Albert, M., Pankovics, P., B Baccarat, H., Pesavento, P. A., Phan, T. G., et al. (2017). Utbrott av neuroinvasivt astrovirus associerat med encefalomyelit, svaghet och förlamning bland avvanda grisar, Ungern. Emerg. Infektera. Dis. 23, 1982–1993. doi: 10.3201 / eid2312. 170804
PubMed Abstrakt / CrossRef fulltext / Google Scholar
han är en av de mest kända och mest kända i världen. (2017). Indikation på överföring mellan arter av astrovirus associerat med encefalit hos får och nötkreatur. Emerg. Infektera. Dis. 23, 1604–1608. doi: 10.3201 / eid2309.170168
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Bouzalas, I. G., W., D., Selimovic-Hamza, S., Dr., C., Bruggmann, R., och Seuberlich, T. (2016). Fullgenombaserad molekylär karakterisering av encefalitassocierade bovinastrovirus. Infektera. Genet. Evol. 44, 162-168. doi: 10.1016/J. meegid.06/052/2016
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Bouzalas, I. G., W., D., Walland, J., Dr., C., Zurbriggen, A., Vandevelde, M., et al. (2014). Neurotropiskt astrovirus hos nötkreatur med nonsuppurativ encefalit i Europa. J. Clin. Mikrobiol. 52, 3318–3324. doi: 10.1128/JCM.01195-14
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Cantile, C. och Youssef, S. (2016). ”Nervsystemet”, i Jubb, Kennedy och Palmers patologi av husdjur, 6: e Edn, ed. M. G. Maxie (Saint Louis, MO: Elsevier), 205-406.
Google Scholar
Deiss, R., Selimovic-Hamza, S., Seuberlich, T. och Meylan, M. (2017). Neurologiska kliniska tecken hos nötkreatur med astrovirusassocierad encefalit. J. Vet. Praktikant. Med. 31, 1209–1214. doi: 10.1111 / jvim.14728
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Donato, C. och Vijaykrishna, D. (2017). Det breda värdområdet och genetisk mångfald av däggdjur och fågel astrovirus. Virus 9: E102. doi: 10.3390 / v9050102
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Drummond, aj, Suchard, ma, Xie, D. och Rambaut, A. (2012). Bayesian fylogenetik med BEAUti och odjuret 1.7. Mol. Biol. Evol. 29, 1969–1973. doi: 10.1093 / molbev / mss075
PubMed Abstrakt / CrossRef fulltext / Google Scholar
han är en av de mest kända och mest kända i världen. (2016). Chlamydia pecorum: foster-och placentalskador vid sporadisk kaprinabort. J. Vet. Diagn. Investera. 28, 184–189. doi: 10.1177/1040638715625729
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Hirashima, Y., Okada, D., Shibata, S., Yoshida, S., Fujisono, S., Omatsu, T., et al. (2018). Hela genomanalys av ett nytt neurotropiskt bovint astrovirus detekterat i en japansk svart steer med icke-suppurativ encefalomyelit i Japan. Båge. Virol. 163, 2805–2810. doi: 10.1007 / s00705-018-3898-3
PubMed Abstrakt / CrossRef fulltext / Google Scholar
internationella utskottet för taxonomi av virus (2018). Master Species List (msl32), Version 1. Tillgänglig på: https://talk.ictvonline.org/files/master-species-lists/m/msl/7185(åtkomst mars, 2019).
Jenkins, G. M., Rambaut, A., Pybus, O. G. Och Holmes, E. C. (2002). Priser för molekylär utveckling i RNA-virus: en kvantitativ fylogenetisk analys. J. Mol. Evol. 54, 156–165. doi: 10.1007 / s00239-001-0064-3
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Kumar, S., Stecher, G. och Tamura, K. (2016). MEGA7: molecular evolutionary genetics analysis version 7.0 för större datamängder. Mol. Biol. Evol. 33, 1870–1874. doi: 10.1093 / molbev / msw054
PubMed Abstrakt / CrossRef fulltext / Google Scholar
li, L., Diab, S., McGraw, S., Barr, B., Traslavina, R., Higgins, R., et al. (2013). Divergerande astrovirus associerat med neurologisk sjukdom hos nötkreatur. Emerg. Infektera. Dis. 19, 1385–1392. doi: 10.3201 / eid1909. 130682
PubMed Abstrakt / CrossRef fulltext / Google Scholar
lole, K. S., Bollinger, R. C., Paranjape, R. S., Gadkari, D., Kulkarni, S. S., Novak, N. G., et al. (1999). Fullängds humant immunbristvirus typ 1 Genom från subtyp C-infekterade serokonverterare i Indien, med bevis på rekombination av intersubtyp. J. Virol. 73, 152–160.
PubMed Abstrakt / Google Scholar
Naccache, S. N. Han är en av de mest kända och mest kända i världen. (2015). Diagnos av neuroinvasiv astrovirusinfektion hos en immunkomprometterad vuxen med encefalit genom opartisk nästa generations sekvensering. Clin. Infektera. Dis. 60, 919–923. doi: 10.1093 / cid / ciu912
PubMed Abstrakt / CrossRef fulltext / Google Scholar
han är en av de mest kända och mest kända i världen. (2011). Patologi och immunhistokemiska fynd av West Nile-virusinfektion i psittaciformes. Veterinär. Pathol. 48, 975–984. doi: 10.1177/0300985810391112
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Pfaff, F., Schlottau, K., Scholes, S., Courtenay, A., Hoffmann, B., H. (2017). Ett nytt astrovirus associerat med encefalit och ganglioneurit hos tamfår. Transbound. Emerg. Dis. 64, 677–682. doi: 10.1111 / tbed.12623
PubMed Abstrakt / CrossRef fulltext / Google Scholar
han var en av de mest kända och mest kända. (2010). Astrovirus encefalit hos pojke med X-länkad agammaglobulinemi. Emerg. Infektera. Dis. 16, 918–925. doi: 10.3201 / eid1606. 091536
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Rambaut, A., Lam, T. T., Max Carvalho, L. och Pybus, O. G. (2016). Utforska den tidsmässiga strukturen hos heterokrona sekvenser med hjälp av TempEst (formely Path-O-Gen). Virus Evol. 2: vew007. doi: 10.1093 / ve / vew007
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Reuter, G., Pankovics, P. och Boros, A. (2018). Nonsuppurativ (aseptisk) meningoencefalomyelit associerad med neurovirulenta astrovirusinfektioner hos människor och djur. Clin. Mikrobiol. Rev. 31, e00040-18. doi: 10.1128 / CMR.00040-18
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Schlottau, K., Schulze, C., Bilk, S., Hanke, D., H Aubbiper, D., öl, M., et al. (2016). Detektion av ett nytt bovint astrovirus hos en ko med encefalit. Transbound. Emerg. Dis. 63, 253–259. doi: 10.1111 / tbed.12493
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Selimovic-Hamza, S., Boujon, C. L., Hilbe, M., Oevermann, A. och Seuberlich, T. (2017a). Frekvens och patologisk fenotyp av bovint astrovirus CH13/NeuroS1-infektion hos nötkreatur med neurologisk sjukdom: mot bedömning av orsakssamband. Virus 9: E12. doi: 10.3390 / v9010012
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Selimovic-Hamza, S., Sanchez, S., Philibert, H., Clark, E. G. och Seuberlich, T. (2017b). Bovin astrovirusinfektion hos feedlot nötkreatur med neurologisk sjukdom i västra Kanada. Kan. Veterinär. J. 58, 601-603.
Google Scholar
Selimovic-Hamza, S., Bouzalas, I. G., Vandevelde, M., Oevermann, A. och Seuberlich, T. (2016). Detektion av astrovirus i historiska fall av europeisk sporadisk bovin encefalit, Schweiz 1958-1976. Front. Veterinär. Sci. 3:91. doi: 10.3389/fvets.2016.00091
PubMed Abstrakt / CrossRef Fulltext / Google Scholar
Seuberlich, T., W., D., Selimovic-Hamza, S., Dr. Han var en av de mest kända. (2016). Identifiering av ett andra encefalitassocierat astrovirus hos nötkreatur. Emerg. Mikrober Infekterar. 5: e5. doi: 10.1038 / emi.2016.5
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Spinato, M. T., Vince, A., Cai, H. och Ojkic, D. (2017). Identifiering av bovint astrovirus i fall av bovin icke-suppurativ encefalit i östra Kanada. Kan. Veterinär. J. 58, 607-609.
Google Scholar
Stein, Lt ,Rech, rr, Harrison, L. och Brown, C. C. (2010). Immunohistokemisk studie av rabiesvirus inom centrala nervsystemet hos inhemska och vilda arter. Veterinär. Pathol. 47, 630–633. doi: 10.1177/0300985810370013
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Trifinopoulos, J., Nguyen, Lt, von Haeseler, A. Och Minh, B. Q. (2016). W-IQ-TREE: ett snabbt online fylogenetiskt verktyg för maximal sannolikhetsanalys. Nukleinsyror Res. 44, W232-W235. doi: 10.1093 / nar / gkw256
PubMed Abstrakt / CrossRef fulltext / Google Scholar
Tse, H., Chan, W. M., Tsoi, H. W. Han är en av de mest kända i världen. (2011). Återupptäckt och genomisk karakterisering av bovint astrovirus. J. Gen. Virol. 92 (Pt 8), 1888-1998. doi: 10.1099 / vir.0.030817-0
PubMed Abstrakt / CrossRef Fulltext / Google Scholar