計算神経科学におけるFrontiers
視覚腹側ストリームのモデルにおけるミュラー-ライアー錯視の誤差を減少させる
複雑な細胞に関する解説
by Zeman,A.,Obst,O.,And Brooks,K.R.(2014年)。 フロント Comput。 ニューロシ… 8:112. ドイ:10.3389/fncom.2014.00112
視覚的錯視は、観察された画像から完全に説明することはできませんが、視覚システムの内部の仕組みから生じる感覚的知覚です。 それらの中で、私たちは画像に物理的に存在しないものを知覚し、神経科学者にとって興味深いのは、私たちが通常認識していない視覚処理を明らか 例えば、同時コントラスト錯視では、絶対値で輝度を知覚するのではなく、視覚システムが周囲との関係で物体の輝度を計算することを理解することができます(図1A)。
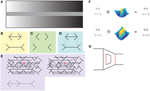
図1. (A)同時コントラスト錯視では、暗い背景がそれを囲むと、均一に灰色の中央のバーが右側に明るく表示されます。 (B)古典的な形のMüller-Lyer錯覚では、矢頭を持つ水平線は矢尾を持つ水平線よりも短く見えます。 (C)錯覚も水平線なしで存在しています。 (D)たとえば、頂点が垂直に整列しているかどうかを判断することによって、視聴者が局所的な特徴を分析するときに錯覚が存在しないことに注意し この認識は、異なる長さを有する水平線の幻想的な効果と矛盾するにもかかわらず、頂点が垂直に整列されることが理解され得る(B)。 (E)連続した錯覚のバージョン(下部に描かれている)は、線の背景内に隠されています。 Müller-Lyerの図は、両方の画像が目の輻輳を減少させることによって融合されるとき、すなわち、画像の平面の後ろに物体を集中させるかのように、背景の上に (F)低レベルの説明では、視覚処理の初期段階でのセンターサラウンド(上のパネル)と単純なセル(下のパネル)のローパス特性から錯覚が生じると述べています。 この仮説は、Zemanらの結果によって支持されなかった。 (G)”carpentered world”の説明では、矢頭と尾は線が異なる深さの角であることを示し、視覚システムはこれを考慮して線のサイズを計算すると述べています。 赤い線の長さは同じです。
私たちの感覚知覚を刺激の物理的特性から解離させることにより、視覚錯覚は神経科学者に私たちの感覚経験の根底にある神経機構を研究す, 2012). 視覚的な錯覚が作成する顕著な知覚は、内部処理から生じるという事実とともに、研究者に錯覚が発生する脳内のメカニズムと場所を探索するよう しかし、幻想は他の知覚現象と同様に説明するのが難しいことが証明されています。
いくつかの幻想の生理学的起源は動物で調査されており、そのうちのいくつかは人間と同様にそれらを知覚することが知られている(Tudusciuc and Nieder、2010)。 本研究は、視床や一次および二次視覚野などの構造における視覚処理の初期段階に、視覚マスキング、フラッシュ抑制、充填、運動誘発深さ、環状知覚(ランダムドットステレオグラム)などの知覚現象が存在することを示している(Carney et al. ら、1 9 8 9;Macknik e t a l. ら、2 0 0 0;von der H Eydt e t a l. ら、2 0 0 0;Grinvald and Hildesheim,2 0 0 4;Wilke e t a l., 2009).
ミュラー-ライアー錯視(MLI)は、古典的な形で、端点に矢頭または矢爪があるかどうかに応じて異なる長さを持つと認識される二つの水平線セグメントからなる、単純で多くの研究されている幾何学的錯視である(図1B–E)。 幻想の背後にある神経機構を理解するための努力でZemanらによる以前の仕事。 (2013)は、mliが霊長類視覚系の多くの特徴を組み込んだモデルである多層人工ネットワークHMAX中に存在することを実証した(Serre et al., 2005). 著者らは、最初に、人間の錯覚を呼び起こさない構成で提示された、短くて長い水平軸の画像を分類するようにネットワークを訓練した。 この訓練の後、彼らはネットワークに、古典的なMLIを含む画像の軸の長さを分類するように依頼しました。
結果は、hmaxネットワークが水平軸の分類に偏りを示し、矢頭を持つものを実際よりも短いものとして分類したことを示しています。 興味深いことに、バイアスの大きさは人間で測定されたものと同様であり、この効果はフィンの角度によっても変調され、角度が小さい(水平軸に近い) 重要なことに、著者らは、最終的な分類層、すなわち画像を長または短として分類する層は、空間周波数の高い単位にのみ依存しないことを実証した。 この結果は、センターサラウンドのローパス特性を示す錯覚の低レベルの説明を支持することができず、単純なセルが錯覚の主な原因である可能性があります(図1F)。 さらに、ネットワークが自然な画像で訓練されておらず、深さに関連する情報が含まれていないことを考えると、錯覚の高レベルの”carpentered world”説明も好まれませんでした(図1G;Segall et al.,1963;Ninio,2014).
Zeman et al. (2014)は、単純な細胞の層によって処理された後に錯覚の大きさが増加し、複雑な細胞の層によって処理された後に減少することを実証することによ 複雑な細胞による錯覚の減少は、位置不変性(その空間的位置にもかかわらず刺激に応答する能力)の特性が、それらのニューロンを錯覚によって誘導されたバイアスに対して感受性を低下させる可能性があることを示唆している。 これらの新しい結果は、MLIの大きさは、異なる神経集団間で異なって表現される可能性があり、画像のより抽象的な表現は、幻想的な効果に敏感ではな
錯覚の背後にあるメカニズムはまだとらえどころのないものです。 Zemanらが示すように、低レベルの説明は、その魅力的な単純さにもかかわらず、完全な話ではないかもしれません。 ランダムドットステレオグラムや他の両眼の錯覚で示されているように(図1E)、mliは、輝度コントラストがなくても、単純な中心サラウンド受容場のものを超えた処理レベルで生成することができる(Julesz、1971)。 “Carpentered world”仮説は錯覚を説明する必要はないが、頭頂および後頭-側頭皮質の関与は、より高い認知過程が関与している可能性が高いことを示唆している(Weidner and Fink,2007;Mancini et al., 2011).
MLIは、直感的に単純な命令”二つの水平線の長さを比較する”が、主観的に感じるほど直接的に視覚システムによって運ばれないことを示しています。 視覚システムが図面全体で何か他のものを比較していることは明らかであり、それはローカル情報ではなく、完全な視覚オブジェクトに関連してい サイズについて尋ねられたとき、私達の視覚システムは完全な目的のサイズを判断しているかもしれません。 これは、例えば、矢印の端点が垂直に整列しているかどうかを判断しようとすることによって、Müller-Lyer図面の局所的な特徴に注意を集中させることによ 頂点が垂直に整列されていることは、図1Bまたは図1Cでさえ、錯覚を示す知覚が局所レベルに存在しないことを理解することができる。
MLI錯視は一見単純な知覚体験であり、神経科学者の注目を集め続けている。 Zemanらの研究は、霊長類のような視覚系内で錯覚を生成するためには、視覚ニューロンのローパスフィルタリング特性と「carpentered world」仮説の2つの錯覚の原因が必 脳が視覚的に識別された物体の大きさを推定し、比較するメカニズムを解明するためには、今後の研究が必要である。
利益相反声明
著者らは、この研究は利益相反の可能性があると解釈される可能性のある商業的または財政的関係がない場合に行われたと宣言
謝辞
我々は、ディレクシオンGeneral de Asuntos del Personal Académico de la Universidad Nacional Autónoma de México及びConsejo Nacional de Ciencia y Tecnologíaからの支援を認識する。 私たちは、技術的支援のためのエドガー Bolañosに感謝します。
Carney,T.,Paradiso,M.A.,And Freeman,R.D.(1989). 猫の皮質ニューロンにおけるPulfrich効果の生理学的相関。 ビジョンRes.29,155-165. 土井: 10.1016/0042-6989(89)90121-1
Pubmed Abstract/Pubmed Full Text|CrossRef Full Text|Google Scholar
Eagleman,D.M.(2001). 視覚錯視と神経生物学。 ナット ニューロシー牧師。 2, 920–926. 土井: 10.1038/35104092
Pubmed Abstract/Pubmed Full Text|CrossRef Full Text|Google Scholar
ることを示した。 VSDI:皮質ダイナミクスの機能イメージングの新しい時代。 ナット ニューロシー牧師。 5, 874–885. doi:10.1038/nrn1536
Pubmed要約/Pubmed全文/CrossRef全文/Google Scholar
Julesz,B.(1971). サイクロプスの知覚の基礎。 オックスフォード:U.Chicago Press。
Macknik,S.L.,Martinez-Conde,S.,And Haglund,M.M.(2000). 可視性と視覚マスキングにおける時空間エッジの役割。 プロク… ナトル アカド サイ… 97,7556-7560 ドイ:10.1073/pnas.110142097
Pubmed要約/Pubmed全文/CrossRef全文|Google Scholar
Mancini,F.,Bolognini,N.,Bricolo,E.,And Vallar,G.(2011). 後頭部-側頭皮質におけるクロスモーダル処理:ミュラー-ライアー錯視のTMS研究。 J.Cogn. ニューロシ… 23, 1987–1997. ドイ:10.1162/jocn.2010.21561
Pubmed要約/Pubmed全文/CrossRef全文|Google Scholar
ニニオ,J.(2014). 幾何学的な幻想は、あなたが彼らがいると思う場所に常にではありません: いくつかの古典的で古典的ではない幻想のレビュー、およびそれらを記述する方法。 フロント ハム ニューロシ… 8:856. ドイ:10.3389/fnhum.2014.00856
Pubmed要約/Pubmed全文/CrossRef全文/Google Scholar
Panagiotaropoulos,T.I.,Deco,G.,Kapoor,V.,And Logothetis,N.K.(2012). ニューロン放電とガンマ振動は、明示的に外側前頭前野の視覚意識を反映しています。 74,924-935 土井:10.1016/j.neuron.2012.04.013
Pubmed要約/Pubmed全文/CrossRef全文/Google Scholar
とができるようになることを期待しています。 幾何学的幻想の認識における文化的違い。 科学139,769-771. 土井:10.1126/科学。139.3556.769
Pubmed要約/Pubmed全文/CrossRef全文|Google Scholar
とができるようになった。 “視覚皮質に触発された特徴を持つ物体認識”IEEE Computer Society Conference on Computer Vision and Pattern Recognition,2005. 2005年、Vol. 2(IEEE),994-1000.
とができます。 サルとヒトにおける長さの判断とミュラー-ライアー錯視の比較。 経験値 脳Res.207,221-231. ドイ:10.1007/s00221-010-2452-7
Pubmed Abstract/Pubmed Full Text|CrossRef Full Text|Google Scholar
von der Heydt,R.,Zhou,H.,And Friedman,H.S.(2000). サルの視覚野における立体エッジの表現。 ビジョンRes.40、1955-1967。 土井:10.1016/0042-6989(00)00044-4
Pubmed Abstract/Pubmed Full Text|CrossRef Full Text|Google Scholar
とができます。 ミュラー-ライアー錯視の根底にある神経機構と視覚空間判断との相互作用。 セレブ 17,878-884 doi:10.1093/cercor/bhk042
Pubmed要約/Pubmed全文/CrossRef全文/Google Scholar
とができるようになることを期待しています。 視床における神経活動は、知覚抑制を反映している。 プロク… ナトル アカド サイ… 106,9465-9470 ドイ:10.1073/pnas.0900714106
Pubmed要約/Pubmed全文/CrossRef全文|Google Scholar
とができるようになりました。 複雑な細胞は、視覚腹側ストリームのモデルにおけるMüller-Lyer錯覚の誤差を減少させる。 フロント Comput。 ニューロシ… 8:112. ドイ:10.3389/fncom.2014.00112
Pubmed要約/Pubmed全文/CrossRef全文/Google Scholar
とができるようになりました。 生物学的物体認識の計算モデルにおけるmüller-lyer錯視。 PLoS ONE8:e56126. doi:10.1371/ジャーナル。ポネ0056126
Pubmed要約|Pubmed全文/CrossRef全文/Google Scholar