Grenser I Pediatri
Bakgrunn
et økende antall studier har vist AT RPS6KA3 er en molekylær etiologi av Coffin–Lowry syndrom (CLS) (1, 2), Et x-bundet semidominant syndrom som først ble rapportert Av Coffin i 1966 og preget av kort statur, ansiktsdysmorfisme, alvorlig til dyp intellektuell funksjonshemming (ID), motorisk utviklingshemming. forsinkelse, Progressive Skjelettdeformiteter, Tannlidelser, Hørselsdefekt Og Annen Medfødt Deformitet Hos menn, Med Intellekt Varierer Fra Normal Til Alvorlig nedsatt i heterozygote kvinner (3-6).
proteinet som er ansvarlig FOR RPS6KA3 er det ribosomale proteinet s6 kinase polypeptid 3 (RSK2), som er en serin/treoninkinase i en familie av mitogenaktiverte proteinkinaser (4). RSK2-proteinet består av to funksjonelle kinase-domener som aktiveres på en sekvensiell måte ved en rekke fosforyleringer, for eksempel PDK-dokkingssted (4). RSK2 har vist seg å være relatert til å spørre celleproliferasjon, blokkere celledifferensiering og beskytte celler fra apoptose (7, 8). Hittil har det vært rapporter om tilfeller med forskjellige symptomer og med over 128 forskjellige mutasjoner i rsk2-genet delt inn i 22 eksoner av chrXp22 .2 (RPS6KA3) (4, 5). Nylig har mutasjonene AV RPS6KA3 vist seg å være relatert til hjertesykdom, osteosarkom, foramen magnum kompresjon, Drop episoder og så videre (7, 9-12).
på Verdensbasis kan den spekulerte prevalensen av CLS være 1/50 000-1 / 100 000 og omtrent 70-80% av probandene har ingen familiehistorie, mens 20-30% har mer enn ett berørt familiemedlem som rapportert (13). Ennå er det ingen kur for denne patologien. Her rapporterer vi en mutasjon AV RPS6KA3 Ved Xp22 i En Kinesisk gutt. Fenotypisk utseende var typisk, inkludert vekst-og utviklingshemming, som beskrevet i tidligere rapporter.
Saksfremstilling
probanden er en 12 måneder gammel gutt med typisk ansiktsdysmorfisme, hørselsdefekt og benabnormalitet (Tabell 1). Han ble født etter en normal graviditet og levert med fødselsvekt på 2,9 kg (10. persentil) og fødselslengde på 45 cm (3.persentil) ved 38 uker, sammenlignet MED WHO Barnevekststandarder i 2006. Ansiktsutseendet presenterer bølgende panne, fremtredende ører, vidt adskilte øyne, ned skrå palpebrale sprekker, kort nese med bred columella, tykk alae nasi og septum, tykk og everted underlip (Figur 1A,B). Løvtennene ble utbrudd ved 8 måneder (ikke forsinket) (Figur 1c). Hendene er korte, kjøttfulle, og med bemerkelsesverdig hyperextensible fingre som taper fra bred til smal med små terminale phalanges og negler (Figurer 1D, 2D). Men det var ingen deformitet av hans foramen magnum eller ryggsøyle (Figur 2B,C). Vekten på 12 måneder er 8,2 kg og høyden er 68.2 cm (< -3.17 z score, WHO). Benmetabolismen og IGF-1Α er forstyrrelse (Vit d 45.2 nmol/l, IGF-1Α < 25 ng/mL). Han begynte å sitte alene på 9 måneder og kunne ikke stå uten hjelp til 12 måneder. Ved 12 måneders alder var hans intelligenskvotient (IQ) 56 i Henhold Til Gesell Developmental Schedules. Han hadde problemer med å bli sittende eller konsentrere seg under oppgavens fullføring. Hans auditive terskel for auditiv hjernestammen respons (ABR) er > 85 db og er diagnostisert som en hørselsforstyrrelse. Den magnetiske resonans imaging (MRI) viste utvidelse av bilaterale ventrikler og mindre cerebral hvit substans (Figur 2a).

Tabell 1. Den typiske fenotypen Av Coffin-Lowry syndrom av proband.
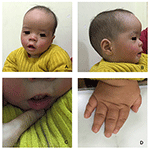
Figur 1. De karakteristiske ansiktsdysmorfismer av En Kiste-Lowry syndrom gutt. Et sidebilde av pasientens ansikt (B), fremtredende ører, vidt adskilte øyne, ned skrå palpebrale sprekker, kort nese med bred columella, tykk alae nasi og septum, tykk og everted underlip (A, C). (D) hendene er korte og med puffy konisk finger, små terminal phalanges og negler.
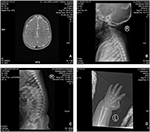
Figur 2. Magnetisk resonans imaging (MRI) og Røntgenbilder Av Coffin-Lowry syndrom saken. (A) MR av cerebral utvikling; (B, C)Cervical og spinalkanalen X-ray fotografier. (D) Nodulær hyperplasi av fingerhaleenden.
for genetisk analyse ble blodprøver oppnådd fra individet. Moren hadde gitt informert samtykke til sine barn. Denne forskningen ble godkjent av bioethics committee for human gene analysis ved Zhejiang University.
neste Generasjons Sekvensering (NGS) analyse ble utført Ved Hjelp Av Agilent Human Genome panel (Agilent Technologies, Inc, Santa Clara, CA, USA). A c.2185 C > T ved chrX20173554 ble påvist i probanden, noe som kan forårsake s.Arg729Trp mutasjon OG rsk2 ustabilitet (Figur 3). Etterfølgende analyse oppdaget ikke samme punktmutasjon i andre familiemedlemmer, inkludert mor, bror (Tilleggsmateriale). Dette indikerte at mikromutasjonen er ny, men ikke arvet fra moren.
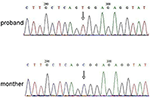
Figur 3. Sanger sekvensering validering for mutasjon AV RPS6KA3 i proband på kromosom Xp22. En mutasjon av c.2185 C > T ble påvist i imprinting kontrollregion fra allel 290-300, som ikke ble påvist i probands mor.
Diskusjon
saken vi presenterer for genetikkevaluering er en 12 måneders gutt som ble født etter en begivenhetsløs graviditet fra friske foreldre. Imidlertid ble hans utviklingsalder forsinket. Som andre berørte CLS-pasienter hadde han den typiske fenotypen observert I CLS, inkludert intellektuell funksjonshemning, retardert motorutvikling, liten statur, tapende fingre, hørselsfeil og karakteristiske ansiktsegenskaper.
RPS6KA3 (OMIM 303600) er kjent for å være mutert hos pasienter med Coffin-Lowry syndrom. Exome c.2185 C > t variant På Xp22 var roman, segregert fra sykdommen, og ble spådd å være skadelig ved hjelp av fem forskjellige i silico programvare (SIFT, Polyphen, MutationTaster, FATHMM, LRT).
den arvede høyden på vår proband er 167,5 cm på grunn av at hans fars høyde er 173 cm mens morens høyde er 150 cm (14). Hvorvidt Den korte staturen Av Coffin-Lowry syndrom kunne behandles med veksthormonanaloger, er fortsatt ikke klart. Rsk er serin / treoninkinaser aktivert AV ERK / MAPK og utgjør fire isoformer (RSK1–4) (15). RSKs fungerer som nedstrøms effektorer AV RTK / Ras / ERK-signalering (Figur 4) (15-17). Som genet som er ansvarlig FOR CLS, SPILLER RSK2 en viktig rolle i celleproliferasjon og migrasjon. Full aktivering AV RSK2 krever fosforylering på flere steder. Vekstfaktorer kan stimulere erk-signalveien til fosforylert RSK2. Som Ramos rapporterte, er erk / RSK2 aktivering involvert i å regulere kreftcelle migrering (18).
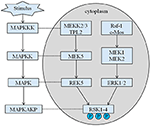
Figur 4. Den foreslåtte veien FOR rsk-relaterte gener. Flere patologiske stimuli, som intrauterin underernæring, utløser nedstrøms signaleringskaskader AV MAPK, som modulerer celleproliferasjon og beinmineralisering.
HVA mer ER, RSK2 ut av funksjon er relatert med beinmineralisering unormal og også induserer ryggraden misdannelser og forkalkninger av ligamentum flavum, som thorax lordose, skoliose, kyfose, og degenerative plate sykdom (19, 20). Tidligere rapporter identifiserte ERK / RSK2 som regulator for beindannelse in vivo (7). Faktisk, Som Marques et al. rapportert, rsk2-mangelfulle mus rekapitulerte det progressive bentapet på grunn av redusert benmineralisering av osteoblaster observert hos pasienter MED CLS (21). Selv om differensieringen av disse cellene in vitro ble drastisk blokkert, ble in vivo fenotypen tydelig tilskrevet en celle autonom reduksjon i osteoblast-aktiviteten i stedet for en reduksjon i tallene deres (7). Interessant NOK ble RSK2 nylig vist å interagere MED TNF-RI til berørte osteoblaster differensiering (18, 22). Som vi vet, kan veksthormon forårsake økt bein mineral tetthet. Fremgangen kan forverre forkalkninger av ligamentum flavum og skjelettdeformitet (23).
Avsluttende Bemerkninger
denne studien er den første som rapporterer Et Kinesisk TILFELLE AV CLS med mutasjon AV RPS6KA3, samt tydelig vekstretardasjon, multiple ansikts abnormiteter, intellektuelle og motoriske funksjonshemninger. Vi spekulerer i at anvendelsen av veksthormonanaloger kan øke forekomsten og invasiviteten av kreft, samt risikoen for ryggradsmisdannelse. Hvis dets effekt og sikkerhet ikke er bekreftet, bør bruk av veksthormonanaloger på PASIENTER AV CLS vurderes spesielt forsiktig. Langsiktige studier med relevant utfall vil være avgjørende i fremtidig klinisk forskning.
Pasientsamtykke
vi fikk informert samtykke for publisering av denne saksrapporten og for bruk av relaterte bilder fra pasientens foreldre.
Forfatterbidrag
JS bidro vesentlig til oppfatningen av dette manuskriptet. YL, LZ, JZ og DW bidro til anskaffelse, analyse eller tolkning av data. YL utarbeidet manuskriptet. JS kritisk revidert manuskriptet for viktig intellektuelt innhold.
Finansiering
dette arbeidet ble støttet av tilskudd fra National Natural Science Foundation Of China (nr. 81501293 og 81773440). Vi erkjenner familien av saken beskrevet i denne rapporten som er klar over og godkjenne denne avidentifiserte konto.
Interessekonflikt
forfatterne erklærer at forskningen ble utført i fravær av kommersielle eller økonomiske forhold som kan tolkes som en potensiell interessekonflikt.
anmelderen BP og håndtering Redaktør erklært sin felles tilknytning på tidspunktet for gjennomgangen.
Supplerende Materiale
Figur S1. Sanger sekvensering validering for mutasjon AV RPS6KA3 i proband bror.
1. Stornetta RL, Zhu JJ. Ras-og Rap-signalering i synaptisk plastisitet og psykiske lidelser. Hjerneforsker (2011) 17: 54-78. doi: 10.1177/1073858410365562
PubMed Abstrakt / Fulltekst / Google Scholar
6. Knish LV. Effekt av vitamin e-preparater brukt i svangerskapet på moderkaken. Pediatr Akus Ginekol. (1966) 2:51–5.
PubMed Abstract | Google Scholar
10. Gschwind M, Foletti G, Baumer A, Bottani A, Novy J. Tilbakevendende nonconvulsive status epilepticus hos en pasient med coffin-lowry syndrom. Mol Syndromol. (2015) 6:91–5. doi: 10.1159/000430429
PubMed Abstrakt / Fulltekst / Google Scholar
12. Martinez HR, Niu MC, Sutton VR, Pignatelli R, Vatta M, Jefferies JL. Coffin-Lowry syndrom og venstre ventrikulær noncompaction kardiomyopati med et restriktivt mønster. AM J Med Genet A (2011) 155A:3030–4. doi: 10.1002 / ajmg.a. 33856
PubMed Abstrakt | CrossRef Fulltekst | Google Scholar
14. Cameron N. GRUNNLEGGENDE programmer for vurdering av skjelett modenhet og prediksjon av voksen høyde Ved Hjelp Av Tanner-Whitehouse metoden. Ann Hum Biol. (1984) 11:261–4. doi: 10.1080/03014468400007151
PubMed Abstrakt / Fulltekst / Google Scholar
15. Romeo Y, Zhang X, Roux PP. Regulering og funksjon AV rsk-familien av proteinkinaser. Biochem J. (2012) 441:553-69. doi: 10.1042 / BJ20110289
PubMed Abstrakt | CrossRef Fulltekst | Google Scholar
17. Beck K, Ehmann N, Andlauer TF, Ljaschenko D, Strecker K, Fischer M, et al. Tap AV Kisten-Lowry syndrom-assosiert genet RSK2 endrer ERK aktivitet, synaptisk funksjon og aksonal transport I Drosophila motoneurons. Dis Modell Mech. (2015) 8:1389–400. doi: 10.1242 / dmm.021246
PubMed Abstrakt | CrossRef Fulltekst | Google Scholar
18. Shi GX, Yang WS, Jin L, Materie ML, Ramos JW. RSK2 driver cellemotilitet ved serinfosforylering AV LARG og aktivering Av Rho Gtpaser. Proc Natl Acad Sci usa. (2018) 115: E190-9. doi: 10.1073 / pnas.1708584115
PubMed Abstrakt / CrossRef Full Tekst / Google Scholar