det humane Amygdaloidkompleks: cellulær arkitektur og dopaminerg innervering
den dopaminerge innervering af det humane AC
AC modtager en betydelig dopaminerg innervering, der hovedsageligt stammer fra A8 -, A9-og A10-ventrale mesencephaliske grupper, og dopamin er en vigtig neurotransmitter i AC, der modulerer indtastningen af information gennem den basolaterale gruppe. Desuden er denne dopaminerge innervering nødvendig til erhvervelse, konsolidering og udryddelse af frygthukommelser såvel som til generering af passende affektive reaktioner, og som tidligere nævnt er dysfunktioner i dette dopaminerge system blevet foreslået som patogene mekanismer i psykiatriske sygdomme såsom schisofreni og stressrelaterede lidelser . Nøjagtige kvantitative data vedrørende mængden af dopaminerge aksoner og deres fordeling i AC fra humane donorer, der ikke var blevet diagnosticeret med neurologiske eller psykiatriske sygdomme før deres død, blev indsamlet af Garca‐Amado og Prensa ved hjælp af DAT immunoreaktivitet som markør for de dopaminerge fibre og stereologiske tilgange. Da iboende dopamininstabilitet forhindrer dets immunodetektion i hjernevæv, der ikke hurtigt er fikseret ved perfusion efter donorens død, havde tidligere undersøgelser, der var fokuseret på at analysere den dopaminerge innervering af den humane AC, brugt TH-proteinet til at detektere dopaminerge profiler. Imidlertid mærker TH protein også noradrenerge og adrenerge fibre i AC . Da AC består af flere nukleare grupper med en lang række sammenkoblinger med hjernebarken, hippocampal dannelse, basale ganglier, thalamus, hypothalamusog hjernestamme (til gennemgang se Refs. ), oplysninger om indholdet af dopaminerge aksoner i hver af de nukleare grupper er nødvendige for bedre at forstå den interne funktionelle organisation af dette kompleks.
den menneskelige AC er målrettet af udbredte DAT‐positive fibre, som er tætte og ujævnt fordelt i hver underafdeling af dette nukleare kompleks (figur 2 og 3). Desuden har deres undersøgelse givet nøjagtige oplysninger om mængden af DAT‐ir-fibre pr. Som vist af disse forfattere varierer mængden af DAT‐ir-aksoner i den menneskelige AC blandt de forskellige kerner i AC og varierer også betydeligt i de forskellige underinddelinger af en given AC-kerne (figur 2), hvilket indikerer funktionelle variationer blandt disse territorier.
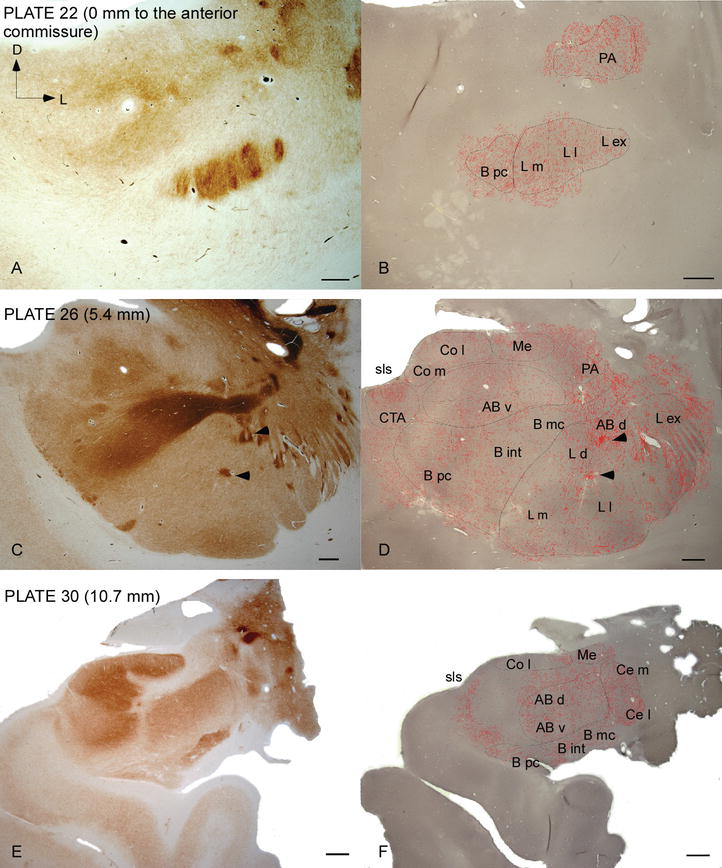
figur 2.
Distribution af DAT‐positive fibre i den humane AC. Serie af to tilstødende koronale sektioner farvet for acetylcholinesterase (AChE) (A, C, E) og DAT (B, D, F) ved tre anteroposterior niveauer af AC, med de tilsvarende plader fra Ref. . Stippling i B, D og F repræsenterer de DAT‐positive aksoner tegnet med camera lucida på 20 liter og overlejret over de samme mikrografer farvet til dat. Pilespidser i C og D angiver pletter med henholdsvis AChE eller dat beriget farvning. For forkortelser se tabel 1. Vægtstang: 1 mm.
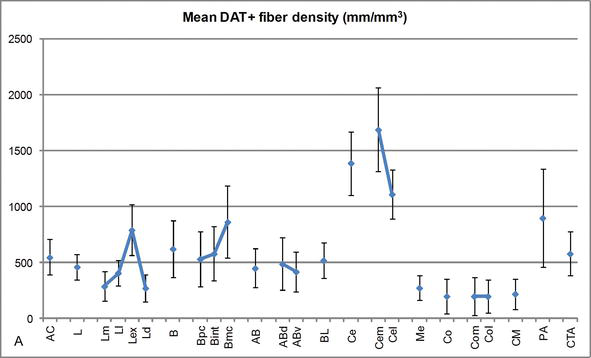
figur 3.
Længdetæthed af DAT‐positive fibre i den humane AC. Gennemsnitlig dat-positiv fiber længde tæthed for hver nuklear gruppe, kerne og nuklear underopdeling af AC. Fejlbjælkerne repræsenterer standardafvigelse. For forkortelser se tabel 1. Modificeret fra Garca-Amado og Prensa .
en af de mest slående gradienter i mængden af DAT‐ir‐fibrene forekom langs den mediolaterale akse af den laterale kerne: den samlede længde af DAT-ir-aksoner spænder fra næsten 300 mm/mm3 i dens mediale underinddeling til næsten 800 mm/mm3 ved dens ydre (mest laterale) underinddeling (figur 2 og 3). Denne store variation i mængden af DAT‐ir-fibre mellem de mediale og laterale sektorer i den laterale kerne kan være relateret til deres differentierede ydre og indre forbindelser. Den laterale kerne ville således være hovedmålet for sensorisk information fra den ydre verden, og den sender tunge fremspring til de andre amygdaloidkerner . Den eksterne underopdeling af den laterale kerne modtager de fleste af disse sensoriske fremspring (figur 2 og 3), og informationen strømmer mod den mediale side af kernen ; derudover har denne AC-region den korteste latenstid af konditionerede reaktioner fremkaldt af sensoriske stimuli forbundet med bivirkninger i følelsesmæssige læringsopgaver . På den anden side modtager den mediale underinddeling af den laterale kerne information fra Højere ordens kortikale behandlingsområder . I hippocampus var DAT-positive aksoner kun til stede i de ydre to tredjedele af det molekylære lag af dentatgyrus, hvor perforeringsvejen slutter , hvilket indikerer, at dopamin potent og selektivt kan regulere input fra entorhinalbarken og dermed de tidlige stadier af hippocampus‐behandling, som det kan være tilfældet for den sensoriske information, der kommer ind i AC-lateral kerne.
den centrale kerne modtager information fra resten af AC-kernerne og er en af AC ‘ s vigtigste outputkerner . Faldende fremspring fra den centrale kerne slutter i en bred mediolateral region af mesencephaliske dopaminceller . Til gengæld modtager denne kerne den tyngste dat-positive dopaminerge innervering af alle AC‐kerner, men dens innervering er ikke ensartet fordelt og falder markant langs en mediolateral gradient, et fund, der stemmer overens med fordelingsmønsteret for TH-ir-fibre . I basal-og tilbehørsbasalkernerne i AC falder indholdet i dat-ir-fibre fra dorsal til ventral sektor, selvom denne gradient er meget mindre markeret i sidstnævnte end i den tidligere kerne (se figur 2a og B) .
reguleringen af ekstracellulære dopaminniveauer styres af forskellige mekanismer i forskellige hjerneområder og er sandsynligvis relateret til dat-indhold. Der henviser således til, at dorsalstriatum og nucleus accumbens viser en” optagelsesdomineret ” regulering (dvs. den mediale præfrontale bark og AC viser en” frigivelsesdomineret ” regulering (dvs .dopamin opretholdes i det ekstracellulære rum mere tid) ; disse fund er enige med observationen om, at der er mere DAT i striatum end i de to andre strukturer.
AC er et hovedmål for mesencephalonfremskrivninger bestående af celler fra substantia nigra pars compacta (A9 dopaminerg gruppe), det ventrale tegmentale område (A10 dopaminerg gruppe) og retrorubral felt (A8 dopaminerg gruppe) . I det humane mesencephalon bugner DAT i neuroner placeret i det laterale ventrale tegmentale område og i substantia nigra pars compacta og er stort set fraværende fra det mediale ventrale tegmentale område . Dat mRNA er mere rigeligt i A9 ventral tier end i dorsal tier . De humane AC-kerner, der indeholder de mest dat‐ir-fibre, svarer til dem, der modtager stærke fremspring fra den ventrale mesencephalon, som også observeret i primater . Der er ikke desto mindre andre AC‐regioner, der viser en høj densitet af DAT-positive fibre, såsom den laterale underinddeling af den centrale kerne, der ikke ser ud til at modtage innervering fra nogen del af den ventrale mellemhjerne . Der er andre mulige kilder til AC-dopamin, der ligger uden for den ventrale midthjerne, men om de bidrager til de DAT‐ir-fibre, der opstår i AC eller ej, er endnu ikke klart. Parabrachialkernen projicerer til AC ‘ s centrale og mediale kerner, og den indeholder formodet dopaminerge neuroner, der ikke bærer dat . Desuden mangler neuronerne i parabrachialkernen, der rager ud til AC, også tyrosinhydroksylase (TH) . Det periakeduktale grå stof er en anden kilde til input til AC, og det indeholder dopaminerge neuroner (dvs .A11-gruppe), der indeholder DAT og projicerer til de centrale og mediale AC-kerner. Denne dopaminerge forbindelse er relevant, da den specifikt er rettet mod den laterale underopdeling af den centrale kerne, et område, der sender efferente fremspring til den mediale underopdeling af den centrale kerne, som igen projicerer tilbage til den periakveduktale grå stofhåndtering “frysning” adfærd hos dyr udsat for en potentielt farlig stimulus . Der er også TH + – celler i den dorsale raphe-kerne , der rager ud til den centrale AC-kerne, men DAT-indholdet af disse celler er endnu ikke bestemt.
den ultrastrukturelle lokalisering af DAT i primat AC er i øjeblikket ukendt. I hjernebarken svarer de fleste af de dat‐mærkede profiler til tynde umyelinerede aksoner, der sjældent danner synapser, hvorimod TH‐mærkede profiler varierer mere i deres diameter, og th‐ir-varicositeter indeholder rigelige vesikler og danner ofte synapser . Derfor, Louis et al. mener, at DAT sandsynligvis vil være begrænset til intervaricose-segmenterne . De specifikke postsynaptiske mål for de dopaminerge fibre, der når den humane AC, er ikke kendt. Flere undersøgelser hos gnavere har vist, at disse fibre fremstiller synapser med både projektionsneuroner og interneuroner . Selvom projektionsneuroner modtager størstedelen af dopaminerge synapser, er CR+ og PV+ interneuron-undermængderne også innerveret af disse fibre, især dem, der indeholder PV . CR + interneuronerne modtager kun 6% af de dopaminerge synapser, mens PV+ – cellerne modtager 40%. I de centrale og basale kerner såvel som i de parakapsulære interkalerede grupper danner de dopaminerge terminaler symmetriske synapser oftere end asymmetriske .
dopaminerge fibre i AC danner perineuronale net omkring Soma af projektionsneuroner og PV+ interneuroner, og 72% af de kontakter, som disse net etablerer med PV+ interneuroner, er synaptiske . Disse net er rigelige i nogle 10-15% af alle PV+ interneuroner, og de ser ud til at undgå andre interneuron-undergrupper. Disse net er funktionelt relateret til den stærke hæmning, der observeres i aktiviteten af projektionsneuronerne i den basolaterale gruppe efter dopaminfrigivelse . Den dopaminerge innervering af de forskellige interneuronpopulationer i AC kunne bidrage til induktion af langsigtede potentieringsmekanismer involveret i konditioneret frygtsamling, hvilket kræver undertrykkelse af Gabaergisk interneuroninhibering af projektionsneuroner . Dopamin hæmmer de” hurtige fyring ” interneuroner , som falder sammen med PV+ interneuroner, og reducerer inhiberingen af projektionsneuroner i den laterale amygdaloidkerne. For nylig, Chu et al. har vist, at dopamin blokerer GABA-frigivelse fra PV+ interneuroner til projektionsneuroner, der virker på type D2 presynaptiske receptorer, men det påvirker ikke frigivelsen af GABA til andre interneurontyper fra denne interneuronpopulation . Blokaden af både D1 og D2 receptorer i den basolaterale gruppe forhindrer frygt betinget erhvervelse .