det mänskliga Amygdaloid-komplexet: cellulär arkitektur och dopaminerg Innervation
den dopaminerga innervationen av den mänskliga AC
AC får en väsentlig dopaminerg innervation med ursprung huvudsakligen från A8, A9 och A10 ventrala mesencefaliska grupper och dopamin är en viktig neurotransmittor i AC som modulerar inmatningen av information genom den basolaterala gruppen. Vidare krävs denna dopaminerga innervation för förvärv, konsolidering och utrotning av rädselminnen samt för att generera lämpliga affektiva svar och, som tidigare nämnts, dysfunktioner i detta dopaminerga system har föreslagits som patogena mekanismer vid psykiatriska sjukdomar såsom schizofreni och stressrelaterade störningar . Exakta kvantitativa data om mängden dopaminerga axoner och deras fördelning i AC från humana givare som inte hade diagnostiserats med neurologiska eller psykiatriska sjukdomar före deras död samlades in av Garcorica‐Amado och Prensa med användning av DAT-immunreaktivitet som markör för dopaminerga fibrer och stereologiska tillvägagångssätt. Eftersom inneboende dopamin instabilitet förhindrar dess immundetektion i hjärnvävnad som inte snabbt har fixats genom perfusion efter givarens död, tidigare studier som var inriktade på att analysera den dopaminerga innervationen av den mänskliga AC hade använt TH-proteinet för att detektera dopaminerga profiler. Emellertid märker TH-proteinet också noradrenerga och adrenerga fibrer i AC . Eftersom AC består av flera kärngrupper med ett brett spektrum av sammankopplingar med hjärnbarken, hippocampalbildning, basala ganglier, thalamus, hypotalamus och hjärnstam (för granskning se Refs. ), information om innehållet av dopaminerga axoner i var och en av kärngrupperna behövs för att bättre förstå den interna funktionella organisationen av detta komplex.
den mänskliga AC riktas mot utbredda DAT‐positiva fibrer, som är täta och ojämnt fördelade i varje underavdelning av detta kärnkomplex (figurerna 2 och 3). Dessutom har deras studie gett korrekt information om mängden DAT-ir-fibrer per neuron i varje amygdaloid territorium. Som framgår av dessa författare varierar mängden dat‐ir-axoner i den mänskliga AC mellan AC: S flera kärnor och varierar också avsevärt i de olika underavdelningarna i en given AC-kärna (Figur 2), vilket indikerar funktionella variationer mellan dessa territorier.
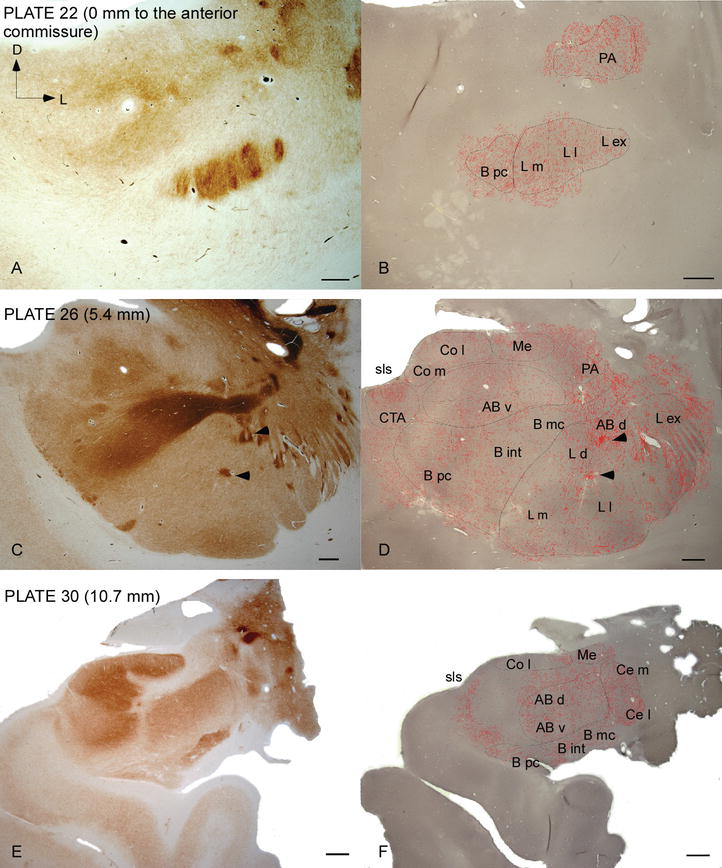
Figur 2.
fördelning av DAT‐positiva fibrer i den mänskliga AC. Serie av två intilliggande koronala sektioner färgade för acetylkolinesteras (AChE) (a, C, E) och DAT (B, D, F) vid tre anteroposterior nivåer av AC, med motsvarande plattor från Ref. . Stipplingen i B, D och F representerar de DAT‐positiva axonerna ritade med camera lucida vid 20 kcal och överlagrade över samma mikrografer färgade för DAT. Pilspetsar i C och D indikerar fläckar med antingen AChE eller DAT-berikad färgning. För förkortningar se Tabell 1. Skala bar: 1 mm.
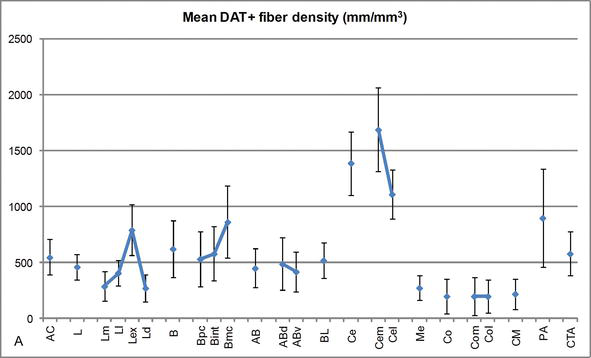
Figur 3.
Längdtäthet av DAT‐positiva fibrer i den mänskliga AC. Genomsnittlig dat-positiv fiberlängdstäthet för varje kärngrupp, kärna och kärnindelning av AC. Felstaplarna representerar standardavvikelse. För förkortningar se Tabell 1. Modifierad från Garcasia-Amado och Prensa .
en av de mest slående gradienterna i mängden DAT‐ir‐fibrer inträffade längs den mediolaterala axeln i den laterala kärnan: den totala längden på DAT-ir-axoner sträcker sig från nästan 300 mm/mm3 i sin mediala underavdelning till nästan 800 mm/mm3 vid dess yttre (mest laterala) underavdelning (figurerna 2 och 3). Denna stora variation i mängden DAT‐ir-fibrer mellan de mediala och laterala sektorerna i den laterala kärnan kan vara relaterad till deras differentierade yttre och inneboende anslutningar. Således skulle sidokärnan vara huvudmålet för sensorisk information från den yttre världen, och den skickar tunga utsprång till de andra amygdaloidkärnorna . Den yttre indelningen av den laterala kärnan mottar de flesta av dessa sensoriska projektioner (figurerna 2 och 3), och informationen strömmar mot den mediala sidan av kärnan ; dessutom har denna AC-region den kortaste latensen av konditionerade svar som framkallas av sensoriska stimuli associerade med biverkningar i emotionella inlärningsuppgifter . Å andra sidan får den mediala indelningen av den laterala kärnan information från kortikala bearbetningsområden med högre ordning . I hippocampus var DAT‐positiva axoner endast närvarande i de yttre två tredjedelarna av molekylskiktet i dentatgyrus, där perforeringsvägen slutar , vilket indikerar att dopamin kan kraftigt och selektivt reglera ingången från entorhinal cortex och därmed de tidiga stadierna av hippocampusbehandling, vilket kan vara fallet för den sensoriska informationen som kommer in i AC‐sidokärnan.
den centrala kärnan tar emot information från resten av AC-kärnorna och är en av de viktigaste utgångskärnorna i AC . Nedåtgående utsprång från den centrala kärnan avslutas i en bred mediolateral region av mesencefaliska dopaminceller . I sin tur mottar denna kärna den tyngsta dat-positiva dopaminerga innervationen av alla AC‐kärnor, men dess innervation är inte jämnt fördelad och minskar markant längs en mediolateral gradient, ett resultat som överensstämmer med fördelningsmönstret för th-ir-fibrer . I de basala och tillhörande basala kärnorna i AC minskar innehållet i DAT‐ir-fibrer från dorsala till ventrala sektorer, även om denna gradient är mycket mindre markerad i den senare än i den tidigare kärnan (se figur 2a och B) .
regleringen av extracellulära dopaminnivåer styrs av distinkta mekanismer i olika hjärnområden och är troligen relaterad till DAT-innehåll. Således, medan dorsalstriatum och nucleus accumbens visar en” upptagningsdominerad ” reglering (dvs. den mediala prefrontala cortexen och AC visar en” frisättningsdominerad ” reglering (dvs dopamin upprätthålls i det extracellulära utrymmet mer tid) ; dessa resultat överensstämmer med observationen att det finns mer DAT i striatum än i de andra två strukturerna .
AC är ett huvudmål för mesencephalonprojektioner som består av celler från substantia nigra pars compacta (A9 dopaminerg grupp), det ventrala tegmentala området (A10 dopaminerg grupp) och det retrorubrala fältet (A8 dopaminerg grupp) . I det mänskliga mesencephalon, dat vimlar i neuroner belägna i det laterala ventrala tegmentala området och i substantia nigra pars compacta och är till stor del frånvarande från det mediala ventrala tegmentala området . DAT mRNA är rikligare i A9 ventral tier än i dorsal tier . De mänskliga AC-kärnorna som innehåller de flesta dat-ir-fibrerna motsvarar de som får starka utsprång från ventral mesencephalon, vilket också observeras i primater . Det finns emellertid andra AC-regioner som visar en hög densitet av DAT-positiva fibrer, såsom den laterala indelningen av den centrala kärnan, som inte verkar ta emot innervation från någon del av den ventrala mellanhjärnan . Det finns andra möjliga källor till AC-dopamin som ligger utanför den ventrala mellanhjärnan, men om de bidrar till DAT‐ir-fibrerna som uppträder i AC eller inte, är det ännu inte klart. Parabrachialkärnan projicerar till de centrala och mediala kärnorna i AC och den innehåller förmodat dopaminerga neuroner som inte bär DAT . Dessutom saknar neuronerna i parabrachialkärnan som projicerar till AC också tyrosinhydroxylas (TH) . Den periaqueductal grå substansen är en annan källa till inmatning till AC och den innehåller dopaminerga neuroner (dvs. A11-grupp) som innehåller DAT och projicerar till de centrala och mediala AC-kärnorna . Denna dopaminerga anslutning är relevant eftersom den specifikt riktar sig mot den laterala indelningen av den centrala kärnan, en region som skickar efferenta utsprång till den mediala indelningen av den centrala kärnan, som i sin tur projicerar tillbaka till periaqueductal grå substans som hanterar ”frysande” beteende hos djur utsatta för en potentiellt farlig stimulans . Det finns också Th+ – celler i dorsal raphe-kärnan som projicerar till den centrala AC-kärnan , men DAT-innehållet i dessa celler har ännu inte fastställts.
den ultrastrukturella lokaliseringen av DAT i primat AC är okänd för närvarande. I hjärnbarken motsvarar de flesta av de DAT‐märkta profilerna tunna omyeliniserade axoner som sällan bildar synapser, medan TH‐märkta profiler varierar mer i diameter och th‐ir-varicositeter innehåller rikliga vesiklar och bildar ofta synapser . Följaktligen, Lewis et al. tror att DAT sannolikt kommer att begränsas till intervaricose-segmenten . De specifika postsynaptiska målen för de dopaminerga fibrerna som når den mänskliga AC är inte kända. Flera studier på gnagare har visat att dessa fibrer gör synapser med både projektionsneuroner och interneuroner . Även om projektionsneuroner får majoriteten av dopaminerga synapser , är CR+ och PV+ interneuron delmängder också innerverade av dessa fibrer, särskilt de som innehåller PV . CR + interneuronerna får endast 6% av de dopaminerga synapserna, medan PV+ – cellerna får 40%. I de centrala och basala kärnorna, liksom i de parakapsulära interkalerade grupperna, bildar de dopaminerga terminalerna symmetriska synapser oftare än asymmetriska .
dopaminerga fibrer i AC-formen perineuronala nät runt Soma av projektionsneuronerna och PV+ interneuronerna, och 72% av kontakterna som dessa nät etablerar med PV+ interneuronerna är synaptiska . Dessa nät är rikliga i cirka 10-15% av alla PV+ interneuroner och de verkar undvika andra interneuron-delmängder. Dessa nät är funktionellt relaterade till den starka inhiberingen som observerats i aktiviteten hos projektionsneuronerna i den basolaterala gruppen efter dopaminfrisättning . Den dopaminerga innervationen av de olika interneuronpopulationerna i AC kan bidra till induktion av långsiktiga potentieringsmekanismer involverade i konditionerat rädsla förvärv, vilket kräver undertryckande av GABAergic interneuronhämning av projektionsneuroner . Dopamin hämmar de” snabba avfyrningarna ” interneuronerna, som sammanfaller med PV+ interneuronerna , och minskar inhiberingen av projektionsneuroner i den laterala amygdaloidkärnan. Mer nyligen, Chu et al. har visat att dopamin blockerar GABA-frisättning från PV + interneuroner till projektionsneuroner som verkar på presynaptiska receptorer av typ D2 men det påverkar inte frisättningen av GABA till andra interneurontyper från denna interneuronpopulation . Blockaden av både D1-och D2-receptorer i den basolaterala gruppen förhindrar rädsla konditionerat förvärv .