ludzki Kompleks migdałowców: Architektura komórkowa i unerwienie dopaminergiczne
unerwienie dopaminergiczne człowieka AC
AC otrzymuje znaczne unerwienie dopaminergiczne pochodzące głównie z brzusznych grup mezencephalicznych A8, A9 i A10, a dopamina jest kluczowym neuroprzekaźnikiem w AC, który moduluje wprowadzanie informacji przez grupę bazolateralną. Ponadto unerwienie dopaminergiczne jest niezbędne do pozyskiwania, utrwalania i wygaszania wspomnień lękowych, jak również do generowania odpowiednich reakcji afektywnych i, jak wspomniano wcześniej, dysfunkcje tego układu dopaminergicznego zostały zaproponowane jako mechanizmy patogenne w chorobach psychiatrycznych, takich jak schizofrenia i zaburzenia związane ze stresem . García‐Amado i Prensa zebrali dokładne dane ilościowe dotyczące ilości aksonów dopaminergicznych i ich dystrybucji w AC od dawców ludzkich, u których przed śmiercią nie zdiagnozowano chorób neurologicznych lub psychiatrycznych, wykorzystując DAT immunoreaktywność jako marker dla włókien dopaminergicznych i podejścia stereologiczne. Ponieważ wewnętrzna niestabilność dopaminy zapobiega jej immunodetekcji w tkance mózgowej, która nie została szybko ustalona przez perfuzję po śmierci dawcy, wcześniejsze badania, które koncentrowały się na analizie unerwienia dopaminergicznego ludzkiego AC, wykorzystywały białko TH do wykrywania profili dopaminergicznych. Jednak białko TH oznacza również włókna noradrenergiczne i adrenergiczne w AC . Ponieważ AC składa się z kilku grup jądrowych z szerokim wachlarzem połączeń z korą mózgową, tworzeniem hipokampa, zwojami podstawnymi, wzgórzem, podwzgórzem i pniem mózgu (w celu przeglądu patrz Refs. ), informacje na temat zawartości aksonów dopaminergicznych w każdej z grup jądrowych są potrzebne do lepszego zrozumienia wewnętrznej organizacji funkcjonalnej tego kompleksu.
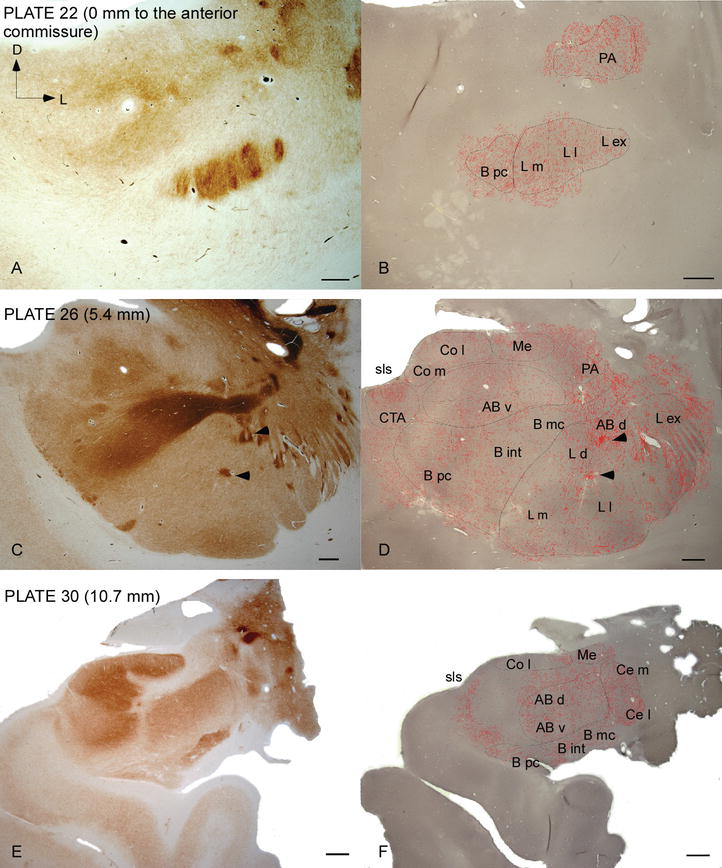
ludzki AC jest celem rozległych włókien DAT-dodatnich, które są gęste i nierównomiernie rozmieszczone w każdym podziale tego kompleksu jądrowego (fig. 2 i 3). Ponadto ich badania dostarczyły dokładnych informacji dotyczących ilości włókien DAT-ir na neuron w każdym obszarze ciała migdałowatego. Jak pokazali ci autorzy, ilość aksonów DAT-ir w ludzkim AC różni się między kilkoma jądrami AC, a także znacznie różni się w różnych poddziałach danego jądra AC (fig. 2), wskazując na różnice funkcjonalne między tymi terytoriami.
Rysunek 2.
Dystrybucja włókien DAT-dodatnich w ludzkim AC. Seria dwóch sąsiednich odcinków koronalnych barwionych na acetylocholinoesterazę (ache) (A, C, E) i DAT (B, D, F) na trzech przednio-tylnych poziomach AC, z odpowiednimi płytkami z Ref. . Punktowanie w B, D i F reprezentuje aksony DAT-dodatnie narysowane za pomocą camera lucida w 20× i nałożone na te same mikrografy barwione dla DAT. Groty strzał w C I d wskazują plamy z albo AChE lub DAT wzbogacony barwienia, odpowiednio. Skróty znajdują się w tabeli 1. Listwa podziałki: 1 mm.
Rysunek 3.
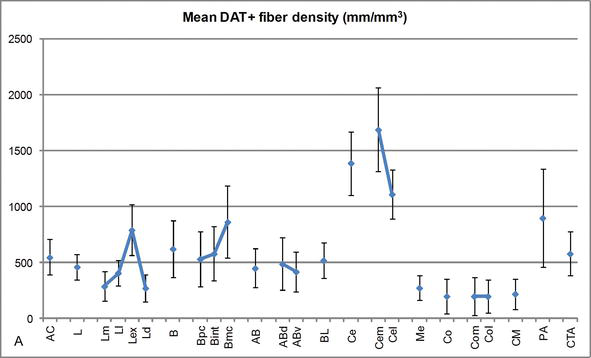
gęstość długości włókien DAT-dodatnich w ludzkim AC. Średnia gęstość długości włókna DAT‐dodatnia dla każdej grupy jądrowej, jądra i podziału jądrowego AC. Paski błędów reprezentują odchylenie standardowe. Skróty znajdują się w tabeli 1. Zmodyfikowany z García-Amado i Prensa .
jeden z najbardziej uderzających gradientów w ilości włókien DAT-ir wystąpił wzdłuż środkowej osi bocznego jądra: całkowita długość aksonów DAT‐IR waha się od prawie 300 mm/mm3 w jego przyśrodkowym podziale do prawie 800 mm/mm3 w jego zewnętrznym (najbardziej bocznym) podziale (fig.2 i 3). Ta duża zmienność w ilości włókien DAT-ir między sektorem przyśrodkowym i bocznym jądra bocznego może być związana z ich zróżnicowanymi połączeniami zewnętrznymi i wewnętrznymi. W ten sposób jądro boczne byłoby głównym celem informacji sensorycznej ze świata zewnętrznego i wysyła ciężkie projekcje do innych jąder amigdaloidów . Zewnętrzny podział bocznego jądra odbiera większość tych projekcji sensorycznych (ryc. 2 i 3), a informacja płynie w kierunku przyśrodkowej strony jądra; ponadto ten region AC ma najkrótsze opóźnienie reakcji uwarunkowanych wywoływanych przez bodźce zmysłowe związane z niekorzystnymi zdarzeniami w emocjonalnych zadaniach uczenia się . Z drugiej strony, przyśrodkowy podział jądra bocznego otrzymuje informacje z obszarów przetwarzania korowego wyższego rzędu . W hipokampie aksony DAT-dodatnie były obecne tylko w zewnętrznych dwóch trzecich warstwy cząsteczkowej zakrętu zębatego, gdzie kończy się szlak perforacji , co wskazuje, że dopamina może silnie i wybiórczo regulować wejście z kory nerwowej, a tym samym wczesne etapy przetwarzania hipokampa, jak to może mieć miejsce w przypadku informacji sensorycznej wprowadzanej do jądra bocznego AC.
jądro Centralne otrzymuje informacje od reszty jąder AC i jest jednym z głównych jąder wyjściowych AC . Zstępujące rzuty z jądra centralnego kończą się w szerokim, śródpiersiowym obszarze komórek mezencefalicznych . Z kolei jądro to otrzymuje najcięższe DAT-dodatnie unerwienie dopaminergiczne ze wszystkich jąder AC, jednak jego unerwienie nie jest równomiernie rozłożone i znacznie zmniejsza się wzdłuż gradientu przyśrodkowo‐bocznego, co jest zgodne ze schematem dystrybucji włókien TH-ir . W jądrach podstawnych i pomocniczych jąder podstawnych AC zawartość włókien DAT‐ir zmniejsza się z sektorów grzbietowych do brzusznych, chociaż gradient ten jest znacznie mniej zaznaczony w tym drugim niż w poprzednim jądrze (patrz rysunek 2a i B).
Regulacja pozakomórkowego poziomu dopaminy jest kontrolowana przez różne mechanizmy w różnych obszarach mózgu i prawdopodobnie jest związana z zawartością DAT. Tak więc, podczas gdy grzbietowe prążkowie i jądro półleżące wykazują regulację „zdominowaną przez wychwyt” (tj. dopamina jest szybko odzyskiwana z przestrzeni zewnątrzkomórkowej, aby zakończyć swoje działanie), przyśrodkowa kora przedczołowa i AC wykazują regulację „zdominowaną przez uwalnianie” (tj. dopamina jest utrzymywana w przestrzeni zewnątrzkomórkowej więcej czasu) ; Wyniki te zgadzają się z obserwacją, że w prążkowiu jest więcej DAT niż w pozostałych dwóch strukturach .
AC jest głównym celem projekcji mezencefalonu złożonego z komórek substancji czarnej Pars compacta (Grupa dopaminergiczna A9), brzusznego obszaru nakrywkowego (Grupa dopaminergiczna A10) i pola retrorubralnego (Grupa dopaminergiczna A8). W ludzkim mezencefalonie DAT obfituje w neurony zlokalizowane w bocznym brzusznym obszarze nakrywki i w substantia nigra Pars compacta i jest w dużej mierze nieobecny w przyśrodkowym brzusznym obszarze nakrywki . DAT mRNA jest bardziej obfite w warstwie brzusznej A9 niż w warstwie grzbietowej . Ludzkie jądra AC, które zawierają najwięcej włókien DAT-ir, odpowiadają tym, które otrzymują silne projekcje z mezencefalonu brzusznego, jak również obserwowane u naczelnych . Istnieją jednak inne regiony AC wykazujące dużą gęstość włókien DAT-dodatnich, takie jak boczny podział jądra centralnego, które nie wydają się otrzymywać unerwienia z żadnej części brzusznego śródmózgowia . Istnieją inne możliwe źródła dopaminy AC, które leżą poza brzusznym śródmózgiem, ale to, czy przyczyniają się do włókien DAT‐ir napotkanych w AC, czy nie, nie jest jeszcze jasne. Jądro parabrachialne wystaje do jądra centralnego i przyśrodkowego AC i zawiera przypuszczalnie neurony dopaminergiczne, które nie przenoszą DAT . Co więcej, neurony jądra parabrachialnego, które projektują AC, również nie mają hydroksylazy tyrozynowej (TH). Periaqueductal szara substancja jest kolejnym źródłem wejścia do AC i zawiera neurony dopaminergiczne (tj. Grupa A11), które zawierają DAT i projekt do centralnego i przyśrodkowego jądra AC . To połączenie dopaminergiczne jest istotne, ponieważ celuje w szczególności w boczny podział jądra centralnego, region, który wysyła eferentne projekcje do przyśrodkowego podziału jądra centralnego, który z kolei rzutuje z powrotem do okołoodbytniczej szarej substancji zajmującej się „zamrażaniem” zachowania u zwierząt narażonych na potencjalnie niebezpieczny bodziec . Istnieją również komórki TH+ w jądrze grzbietowym, które wystają do centralnego jądra AC, ale zawartość DAT w tych komórkach nie została jeszcze określona.
lokalizacja ULTRASTRUKTURALNA DAT u Prymasa AC jest obecnie nieznana. W korze mózgowej większość profili oznaczonych DAT odpowiada cienkim niemielinizowanym aksonom, które rzadko tworzą synapsy, podczas gdy profile znakowane TH różnią się bardziej średnicą, a żylaki TH-ir zawierają obfite pęcherzyki i często tworzą synapsy . W związku z tym Lewis et al. przypuszcza się, że DAT może być ograniczony do segmentów interwarikozy . Specyficzne cele postsynaptyczne włókien dopaminergicznych, które docierają do ludzkiego AC, nie są znane. Kilka badań na gryzoniach wykazało, że włókna te tworzą synapsy zarówno z neuronami projekcyjnymi, jak i interneuronami . Chociaż neurony projekcyjne otrzymują większość synaps dopaminergicznych, podgrupy interneuronów CR+ i PV+ są również unerwione przez te włókna, zwłaszcza te zawierające PV . Interneurony CR+ otrzymują tylko 6% synaps dopaminergicznych, podczas gdy komórki PV+ otrzymują 40% . W jądrze centralnym i podstawnym, a także w parakapsularnych grupach interkalowanych, terminale dopaminergiczne częściej tworzą symetryczne synapsy niż asymetryczne .
włókna dopaminergiczne W AC tworzą siatki krocza wokół soma neuronów projekcyjnych i interneuronów PV+, a 72% kontaktów, które te sieci nawiązują z interneuronami PV+, jest synaptycznych . Sieci te są obfite w około 10-15% wszystkich interneuronów PV+ i wydają się unikać innych podzbiorów interneuronów. Sieci te są funkcjonalnie związane z silnym hamowaniem obserwowanym w aktywności neuronów projekcyjnych grupy zasadowej po uwolnieniu dopaminy . Unerwienie dopaminergiczne różnych populacji interneuronów AC może przyczynić się do indukcji długoterminowych mechanizmów wzmocnienia zaangażowanych w uwarunkowane nabywanie strachu, co wymaga tłumienia Gabaergicznego hamowania interneuronu neuronów projekcyjnych . Dopamina hamuje” szybkie wypalanie ” interneuronów, które pokrywają się z interneuronami PV+ i zmniejsza hamowanie neuronów projekcyjnych w bocznym jądrze migdałowatym. Ostatnio, Chu et al. wykazano, że dopamina blokuje uwalnianie GABA z interneuronów PV+ do neuronów projekcyjnych działających na receptory presynaptyczne typu D2, ale nie wpływa na uwalnianie GABA do innych typów interneuronów z tej populacji interneuronów . Blokada zarówno receptorów D1, jak i D2 w grupie bazolateralnej zapobiega przejawowi lęku.