ihmisen Amygdaloidikompleksi: Cellular Architecture and dopaminerginen innervaatio
ihmisen dopaminerginen innervaatio
AC saa huomattavan dopaminergisen innervaation, joka on peräisin pääasiassa A8 -, A9-ja A10-ventraalisista mesenkefalisista ryhmistä, ja dopamiini on AC: n keskeinen välittäjäaine, joka moduloi informaation tuloa basolateraalisen ryhmän kautta. Lisäksi tätä dopaminergistä innervaatiota tarvitaan pelkomuistien hankkimiseen, vahvistamiseen ja häviämiseen sekä asianmukaisten affektiivisten vasteiden tuottamiseen, ja kuten aiemmin mainittiin, tämän dopaminergisen järjestelmän toimintahäiriöitä on ehdotettu patogeenisiksi mekanismeiksi psykiatrisissa sairauksissa, kuten skitsofreniassa ja stressiin liittyvissä häiriöissä . García‐Amado ja Prensa keräsivät tarkkoja määrällisiä tietoja dopaminergisten aksonien määrästä ja niiden jakautumisesta AC: ssä ihmisluovuttajilta, joilla ei ollut diagnosoitu neurologisia tai psykiatrisia sairauksia ennen kuolemaansa, käyttäen dat‐immunoreaktiivisuutta dopaminergisten kuitujen ja stereologisten lähestymistapojen merkkiaineena. Koska luontainen dopamiinin epävakaus estää sen immunodetektion aivokudoksessa, jota ei ole nopeasti korjattu perfuusiolla luovuttajan kuoleman jälkeen, aiemmissa tutkimuksissa, jotka keskittyivät analysoimaan ihmisen AC: n dopaminergistä innervaatiota, TH-proteiinia oli käytetty dopaminergisten profiilien havaitsemiseen. TH-proteiini merkitsee kuitenkin myös AC: n noradrenergisiä ja adrenergisiä kuituja . Koska AC koostuu useista ydinryhmistä, joilla on laaja valikoima yhteyksiä aivokuoren, hippokampal muodostumista, tyvitumakkeiden, talamus, hypotalamus, ja brainstem (tarkistettavaksi Katso viite. ), tietoa dopaminergisten aksonien sisällöstä kussakin ydinryhmässä tarvitaan tämän kompleksin sisäisen toiminnallisen organisaation ymmärtämiseksi paremmin.
ihmisen AC: n kohteena ovat laajalle levinneet DAT‐positiiviset kuidut, jotka ovat tiheitä ja epätasaisesti jakautuneita tämän ydinkompleksin jokaisessa alaryhmässä (Kuvat 2 ja 3). Lisäksi heidän tutkimuksensa on tuottanut tarkkaa tietoa DAT-ir-kuitujen määrästä hermosolua kohti kullakin mantelitumakkeiden alueella. Kuten nämä kirjoittajat osoittavat, DAT‐ir aksonien määrä ihmisen AC vaihtelee eri ytimissä AC ja myös vaihtelee huomattavasti eri osa tietyn AC tuman (kuva 2), mikä osoittaa toiminnallisia vaihteluita näiden alueiden välillä.
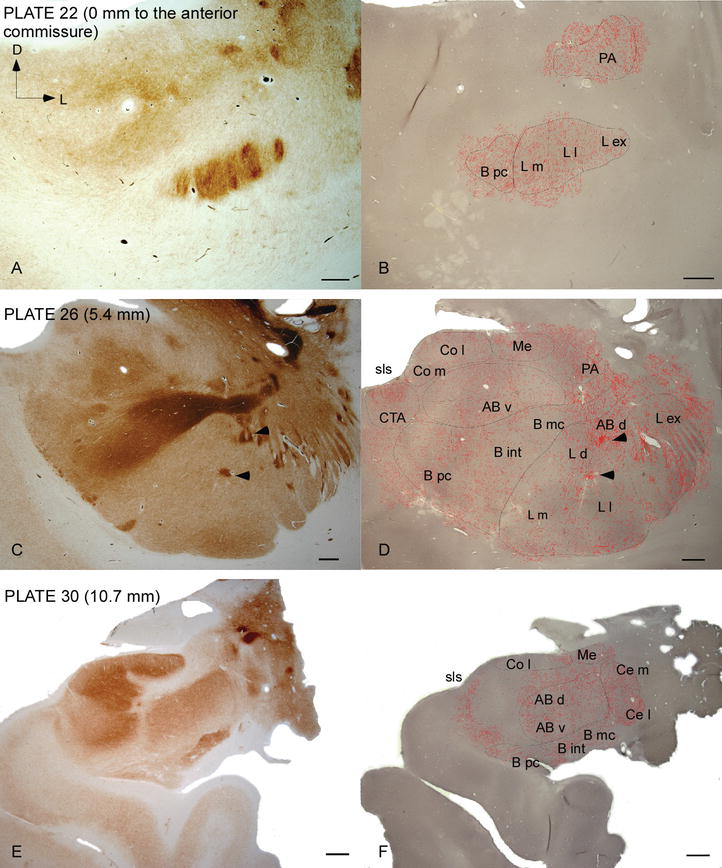
kuva 2.
dat‐positiivisten kuitujen jakautuminen ihmisen AC. Sarja kahden vierekkäisen koronan osat värjätään asetyylikoliiniesteraasi (AChE) (A, C, E) ja DAT (B, D, F) kolmella anteroposterior tasolla AC, vastaavat levyt Ref. . Stippling B, D ja F edustaa dat positiivinen aksonit piirretään kamera lucida 20× ja päällekkäin samojen mikrografit värjätään DAT. Nuolenkärjet C ja D osoittavat laastarit joko AChE tai DAT rikastettu värjäys, vastaavasti. Lyhenteet KS. Taulukko 1. Mittakaari: 1 mm.
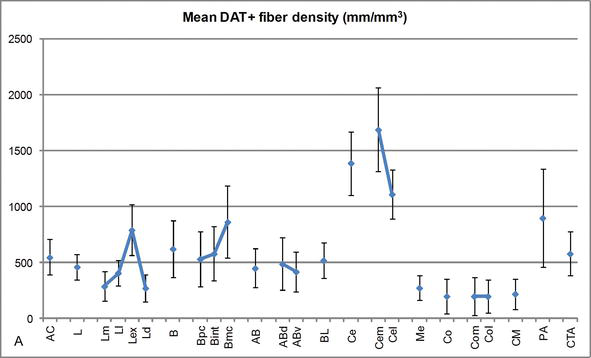
kuva 3.
dat‐positiivisten kuitujen Pituustiheys ihmisen AC: ssä. Keskimääräinen dat-positiivinen kuidun pituuden tiheys jokaiselle YDINRYHMÄLLE, ytimelle ja ydinosastolle AC. Virhepalkit edustavat keskihajontaa. Lyhenteet KS. Taulukko 1. Muokattu García-Amadosta ja Prensasta .
yksi silmiinpistävimmistä kaltevuuksista dat‐ir-kuitujen määrässä tapahtui lateraaliytimen mediolateraalisella akselilla: dat‐ir-aksonien kokonaispituus vaihtelee mediaalisessa osastoinnissa lähes 300 mm/mm3 ja ulkoisessa (sivusuuntaisimmassa) osastossa lähes 800 mm/mm3 (Kuvat 2 ja 3). Tämä suuri vaihtelu dat‐ir-kuitujen määrässä lateraalisen ytimen mediaalisten ja lateraalisten sektoreiden välillä saattaa liittyä niiden erilaistuneisiin ulkoisiin ja sisäisiin yhteyksiin. Lateraalinen Tuma olisi siis ulkoisesta maailmasta tulevan aistinvaraisen informaation pääkohde, ja se lähettää raskaita ulokkeita muille amygdaloidiytimille . Lateraaliytimen ulkoinen osa-alue saa suurimman osan näistä aistiprojektioista (Kuvat 2 ja 3), ja tieto virtaa kohti tuman mediaalista puolta .lisäksi tällä AC-alueella on lyhin latenssi ehdollistuneista reaktioista, joita emotionaalisissa oppimistehtävissä esiintyviin haitallisiin tapahtumiin liittyvät aistiärsykkeet saavat aikaan. Toisaalta lateraaliytimen mediaalinen osastoiminen vastaanottaa tietoa korkeammantasoisilta aivokuoren käsittelyalueilta . Hippokampuksessa DAT-positiivisia aksoneja oli vain hammasproteesigyruksen molekyylikerroksen uloimmassa kahdessa kolmasosassa , jossa perforointireitti päättyy, mikä viittaa siihen, että dopamiini voi voimakkaasti ja selektiivisesti säädellä entorhinaalisesta aivokuoresta tulevaa syöttöä ja siten hippokampuksen käsittelyn alkuvaiheita, kuten AC: n lateraaliytimeen tulevan aistitiedon tapauksessa.
keskusydin vastaanottaa tietoa muilta VAIHTOVIRTAYTIMILTÄ ja on yksi VAIHTOVIRTAYTIMEN tärkeimmistä ulostuloytimistä . Keskusytimestä laskeutuvat ulokkeet päättyvät mesenkefalisten dopamiinisolujen laajaan mediolateraaliseen alueeseen . Tämä ydin puolestaan saa raskaimman dat-positiivisen dopaminergisen innervaation kaikista AC-ytimistä, mutta sen innervaatio ei jakaudu tasaisesti ja pienenee huomattavasti mediolateraalista gradienttia pitkin, mikä on sopusoinnussa TH‐ir-kuitujen jakautumisen kanssa . AC: n tyvi‐ja lisälaitteiden tyviytimissä dat-ir-kuitujen pitoisuus vähenee dorsaalisista ventraalisiin sektoreihin, vaikka tämä gradientti on paljon vähemmän merkitty jälkimmäisessä kuin edellisessä tumassa (KS .Kuva 2A ja B).
solunulkoisen dopamiinipitoisuuden säätelyä säätelevät eri aivoalueiden erilliset mekanismit, ja se liittyy todennäköisesti DAT-pitoisuuteen. Näin ollen, kun taas dorsal striatum ja Tuma accumbens osoittavat ”kertymävaltaista” säätelyä (ts. yksi, jossa dopamiini otetaan nopeasti takaisin solunulkoisesta tilasta sen toiminnan lopettamiseksi), mediaalinen etuotsalohko ja AC osoittavat ”vapautumisvaltaista” säätelyä (ts .dopamiini säilyy solunulkoisessa tilassa enemmän aikaa); nämä havainnot ovat yhtäpitäviä havainnon kanssa, että striatumissa on enemmän DAT: tä kuin kahdessa muussa rakenteessa.
AC on mesenkefaloniprojektioiden pääkohde, joka koostuu substantia nigra Pars compacta-bakteerin (A9-dopaminerginen ryhmä), ventraalisen tegmenttialueen (A10-dopaminerginen ryhmä) ja retrorubaalisen kentän (A8-dopaminerginen ryhmä) soluista . Ihmisen mesencephalon, DAT runsaasti neuronien sijaitsee lateral ventral tegmental alueella ja substantia nigra pars compacta ja on suurelta osin poissa medial ventral tegmental alueella . DAT mRNA on runsaampi A9 ventraalitasossa kuin dorsaalitasossa . Ihmisen AC‐ytimet, jotka sisältävät eniten DAT-ir-kuituja, vastaavat niitä, jotka saavat voimakkaita ulokkeita ventraalisesta mesenkefalonista, kuten on havaittu myös kädellisillä . On kuitenkin olemassa muita AC-alueita, joissa esiintyy runsaasti dat-positiivisia kuituja, kuten keskimmäisen tuman lateraalinen osastoiminen, jotka eivät näytä saavan innervaatiota mistään ventraalisen keskiaivon osasta . On olemassa muita mahdollisia AC dopamiinin lähteitä, jotka sijaitsevat ventraalisen keskiaivon ulkopuolella, mutta onko ne edistävät dat‐ir-kuituja AC: ssä vai ei, ei ole vielä selvää. Parabrakiaalinen ydin projisoituu AC: n keski-ja mediaalisiin ytimiin ja se sisältää oletetusti dopaminergisiä neuroneja, jotka eivät kanna DAT: tä . Lisäksi PARABRAKIAALISEN ytimen neuroneista, jotka projisoituvat AC: hen, puuttuu myös tyrosiinihydroksylaasi (TH) . Periaqueduktaalinen harmaa aine on toinen tulolähde AC: hen ja se sisältää dopaminergisiä neuroneja (eli A11-ryhmää), jotka sisältävät DAT: tä ja projisoivat keskus-ja mediaalisiin AC-ytimiin . Tämä dopaminerginen yhteys on merkityksellinen, koska se kohdistuu erityisesti keskiytimen lateraaliseen osastoimiseen, alueeseen, joka lähettää efferentteja projektioita keskusytimen mediaaliseen osastoimiseen, joka puolestaan projisoi takaisin periaqueduktaalisen harmaan aineen käsittelyn ”jäätymisen” käyttäytymiseen eläimillä, jotka altistuvat mahdollisesti vaaralliselle ärsykkeelle . Dorsaalisessa raphe-tumassa on myös TH+ – soluja , jotka projisoituvat keskimmäiseen AC-tumaan, mutta näiden solujen DAT-pitoisuutta ei ole vielä määritetty.
DAT: n ultrarakenteellista sijaintia KÄDELLISESSÄ AC: ssa ei toistaiseksi tunneta. Vuonna aivokuori, useimmat dat-merkitty profiilit vastaavat ohut unmyelinated aksonit, jotka harvoin muodostavat synapseja, kun taas TH-merkitty profiilit vaihtelevat enemmän niiden halkaisija ja th-ir varicosities sisältää runsaasti rakkuloita ja usein muodostavat synapseja . Näin ollen Lewis et al. uskovat, että DAT on todennäköisesti rajoitettu intervaricose segmentit . Dopaminergisten kuitujen erityisiä postsynaptisia kohteita, jotka saavuttavat ihmisen AC: n, ei tunneta. Useat jyrsijöillä tehdyt tutkimukset ovat osoittaneet, että nämä kuidut muodostavat synapseja, joissa on sekä projektioneuroneja että interneuroneja . Vaikka projisointisolut vastaanottavat valtaosan dopaminergisistä synapseista, CR+ – ja PV+ – interneuron-osajoukot ovat myös näiden kuitujen, erityisesti PV: tä sisältävien, inervoimia . CR+ – interneuronit saavat vain 6% dopaminergisistä synapseista, kun taas PV+ – solut saavat 40% . Keski-ja tyviytimissä sekä parakapsulaarisissa interkaloituneissa ryhmissä dopaminergiset terminaalit muodostavat symmetrisiä synapseja useammin kuin epäsymmetriset .
AC: n dopaminergiset kuidut muodostavat perineuronaalisia verkkoja projektioneuronien Soman ja PV+ interneuronien ympärille, ja 72% näiden verkkojen yhteyksistä PV+ interneuronien kanssa ovat synaptisia . Näitä verkkoja on runsaasti noin 10-15% kaikista PV+ interneuroneista ja ne näyttävät välttävän muita interneuron osajoukkoja. Nämä verkot liittyvät toiminnallisesti basolateraalisen ryhmän projektioneuronien aktiivisuudessa havaittuun voimakkaaseen estoon dopamiinin vapautumisen jälkeen . AC: n eri interneuronipopulaatioiden dopaminerginen innervaatio voi edistää ehdollistuneeseen pelon hankintaan liittyvien pitkäaikaisten potentiaatiomekanismien induktiota, mikä edellyttää projektioneuronien gabaergisen interneuronin eston estämistä . Dopamiini estää ”nopeasti ampuvia” interneuroneja, jotka ovat yhtäpitäviä PV+ – interneuronien kanssa , ja vähentää projektioneuronien estoa lateraalisessa amygdaloidiytimessä. Viime aikoina Chu et al. on osoitettu, että dopamiini estää GABA: n vapautumisen PV+ – interneuroneista tyypin D2 presynaptisiin reseptoreihin vaikuttaviin projisoituihin neuroneihin, mutta se ei vaikuta GABA: n vapautumiseen muihin interneuronityyppeihin tästä interneuronipopulaatiosta . Basolateraalisen ryhmän sekä D1-että D2-reseptorien salpaus estää pelon ehdollistumisen .