a humán Amygdaloid komplex: Sejtstruktúra és dopaminerg beidegzés
a humán AC dopaminerg beidegzése
az AC jelentős dopaminerg beidegzést kap, amely elsősorban az A8, A9 és A10 ventrális mesencephalikus csoportokból származik, és a dopamin kulcsfontosságú neurotranszmitter az AC-ben, amely modulálja az információ bevitelét a bazolaterális csoporton keresztül. Továbbá ez a dopaminerg beidegzés szükséges a félelem emlékeinek megszerzéséhez, konszolidációjához és kihalásához, valamint a megfelelő affektív válaszok létrehozásához, és amint azt korábban említettük, ennek a dopaminerg rendszernek a diszfunkcióit javasolták patogén mechanizmusként pszichiátriai betegségekben, mint például a skizofrénia és a stresszel kapcsolatos rendellenességek . A dopaminerg axonok mennyiségére és az AC-ben való eloszlására vonatkozó pontos kvantitatív adatokat olyan emberi donoroktól gyűjtötték össze, akiknél haláluk előtt nem diagnosztizáltak neurológiai vagy pszichiátriai betegségeket, a Garc Aponca-Amado és a Prensa a DAT immunreaktivitást használva a dopaminerg rostok markereként és sztereológiai megközelítésekkel. Mivel a belső dopamin instabilitás megakadályozza annak immundetektálását az agyszövetben, amelyet a donor halála után nem sikerült gyorsan rögzíteni perfúzióval, a humán AC dopaminerg beidegzésének elemzésére összpontosító korábbi tanulmányok a th fehérjét használták a dopaminerg profilok kimutatására. Azonban a th fehérje a noradrenerg és adrenerg szálakat is jelöli az AC-ben . Mivel az AC több nukleáris csoportból áll, amelyek hatalmas összeköttetésekkel rendelkeznek az agykéreggel, a hippokampusz kialakulásával, a bazális ganglionokkal, a talamusszal, a hipotalamusszal és az agytörzsvel (lásd A Refs-t. ), információra van szükség a dopaminerg axonok tartalmáról az egyes nukleáris csoportokban, hogy jobban megértsük ennek a komplexnek a belső funkcionális szervezetét.
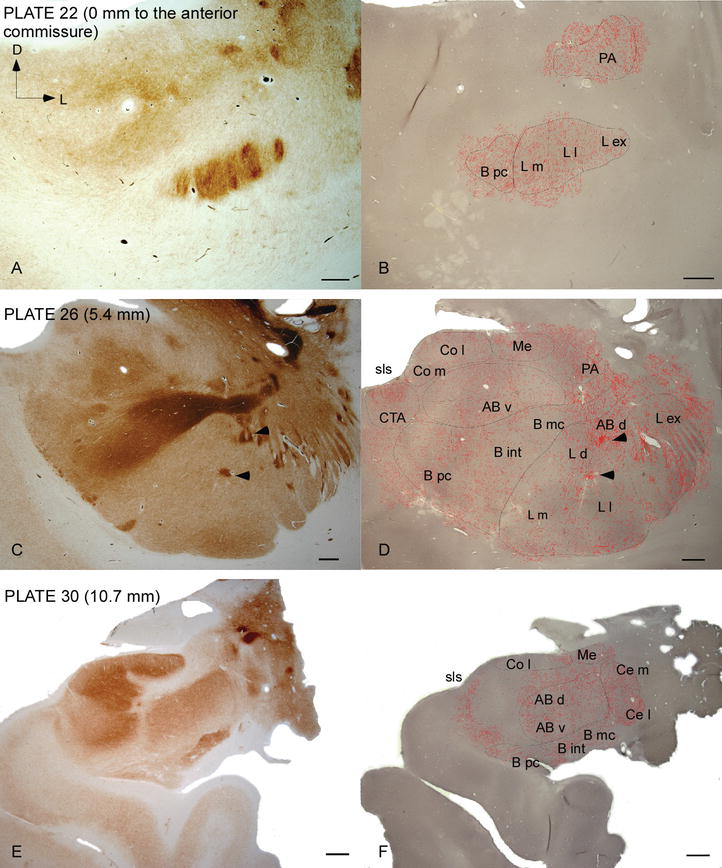
az emberi AC‐t széles körben elterjedt DAT-pozitív szálak célozzák meg, amelyek sűrűek és egyenlőtlenül oszlanak el ennek a nukleáris komplexnek minden alrészében (2.és 3. ábra). Továbbá tanulmányuk pontos információkat adott a DAT-ir szálak mennyiségéről neurononként az egyes amygdaloid területeken. Amint azt ezek a szerzők mutatják, a DAT‐ir axonok mennyisége az emberi AC-ben az AC több magja között változik, és jelentősen változik egy adott AC-mag különböző alosztályaiban is (2.ábra), jelezve a funkcionális eltéréseket ezen területek között.
2. ábra.
dat‐pozitív szálak eloszlása az emberi AC-ben. Az AC három anteroposterior szintjén az acetilkolinészteráz (AChE) (A, C, E) és a DAT (B, D, F) számára festett két szomszédos koronális szakasz sorozata, a megfelelő lemezekkel Ref. . A B, D és F pontozás a camera lucida‐val rajzolt dat-pozitív axonokat jelöli 20 6connig, és ugyanazon a DAT-ra festett mikrográf felett helyezkedik el. A C és D nyílhegyek ache vagy DAT dúsított foltokkal rendelkező foltokat jeleznek, illetőleg. A rövidítéseket Lásd az 1. táblázatban. Méretarány: 1 mm.
3. ábra.
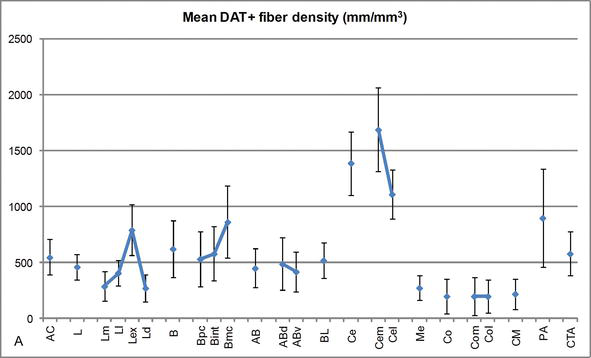
a DAT‐pozitív szálak Hosszsűrűsége az emberi AC-ben. Átlagos dat-pozitív szálhossz-sűrűség az AC minden nukleáris csoportjára, magjára és nukleáris felosztására. A hibasávok a szórást jelentik. A rövidítéseket Lásd az 1. táblázatban. Módosítva Garc! – Amado és Prensa.
az egyik legszembetűnőbb gradiens a DAT‐ir szálak mennyiségében az oldalsó mag mediolaterális tengelye mentén következett be: a DAT‐ir axonok teljes hossza medialis felosztásában közel 300 mm/mm3-től közel 800 mm/mm3-ig terjed a külső (leginkább laterális) felosztásánál (2.és 3. ábra). Ez a nagy eltérés a DAT‐ir rostok mennyiségében a laterális mag mediális és laterális szektorai között összefügghet a differenciált külső és belső kapcsolataikkal. Így az oldalsó mag lenne a külső világból származó szenzoros információk fő célpontja, és nehéz vetületeket küld a többi amygdaloid magnak . A laterális mag külső felosztása kapja a legtöbb ilyen szenzoros vetületet (2 .és 3. ábra), és az információ a mag mediális oldala felé áramlik ; ezenkívül ez az AC régió rendelkezik a legrövidebb késleltetéssel az érzelmi tanulási feladatok során nemkívánatos eseményekhez kapcsolódó szenzoros ingerek által kiváltott kondicionált válaszokkal. Másrészt az oldalsó mag mediális felosztása információkat kap a magasabb rendű kortikális feldolgozási területekről . A hippocampusban a DAT-pozitív axonok csak a dentate gyrus molekuláris rétegének külső kétharmadában voltak jelen , ahol a perforáns út véget ér, jelezve, hogy a dopamin hatékonyan és szelektíven szabályozhatja az entorhinalis kéregből érkező bemenetet és így a hippocampus feldolgozásának korai szakaszát, mint például az AC laterális magba belépő szenzoros információ esetében.
a központi mag információt kap a többi AC magtól, és az AC egyik fő kimeneti magja . A központi magból származó csökkenő vetületek a mesencephalic dopamin sejtek széles mediolaterális régiójában végződnek . Viszont ez a mag kapja az összes AC mag legnehezebb dat-pozitív dopaminerg beidegzését, azonban beidegzése nem egyenletesen oszlik el, és jelentősen csökken a mediolaterális gradiens mentén, ami megegyezik a th‐ir rostok eloszlási mintázatával . Az AC bazális és kiegészítő bazális magjaiban a DAT‐ir rostok tartalma a dorzálisról a ventrális szektorra csökken, bár ez a gradiens sokkal kevésbé markáns az utóbbiban, mint az előbbi magban (lásd a 2a .és B. ábrát).
az extracelluláris dopaminszint szabályozását különböző agyi területek különböző mechanizmusai szabályozzák, és valószínűleg a DAT-tartalommal függ össze. Így, míg a dorsalis striatum és a nucleus accumbens “felvétel‐dominált” szabályozást mutat (pl. az egyikben a dopamin gyorsan visszaáll az extracelluláris térből, hogy befejezze működését), a mediális prefrontális kéreg és az AC “felszabadulás által dominált” szabályozást mutat (azaz a dopamin az extracelluláris térben több időt tart fenn) ; ezek az eredmények egyetértenek azzal a megfigyeléssel, hogy a striatumban több DAT van, mint a másik két struktúrában .
az AC a substantia nigra pars compacta (A9 dopaminerg csoport), a ventrális tegmentális terület (A10 dopaminerg csoport) és a retrorubral mező (A8 dopaminerg csoport) sejtjeiből álló mesencephalon vetületek fő célpontja . Az emberi mesencephalonban a DAT bővelkedik a laterális ventrális tegmentális területen és a substantia nigra pars compacta-ban található neuronokban, és nagyrészt hiányzik a mediális ventrális tegmentális területről . A DAT mRNS bőségesebb az A9 ventrális rétegben, mint a hátsó rétegben . A legtöbb DAT‐ir rostot tartalmazó emberi AC magok megfelelnek azoknak, amelyek erős vetületeket kapnak a ventrális mesencephalonból, amint azt a főemlősöknél is megfigyelték . Vannak azonban más AC régiók, amelyek nagy dat‐pozitív rostok sűrűségét mutatják, például a központi mag oldalirányú felosztása, amelyek úgy tűnik, hogy nem kapnak beidegzést a ventrális középagy bármely részéből . Vannak más lehetséges AC dopaminforrások is, amelyek a ventrális középagyon kívül helyezkednek el, de még nem világos, hogy hozzájárulnak‐e az AC-ben előforduló DAT-ir rostokhoz vagy sem. A parabrachiális mag az AC központi és mediális magjaiba vetül ki, és feltételezhetően dopaminerg neuronokat tartalmaz, amelyek nem hordoznak DAT-ot . Ezenkívül a PARABRACHIÁLIS mag neuronjai, amelyek az AC-re vetülnek, szintén hiányoznak tirozin-hidroxiláz (TH). A periaqueductal gray anyag egy másik bemeneti forrás az AC-hez, és dopaminerg neuronokat (azaz A11 csoportot) tartalmaz, amelyek DAT-ot tartalmaznak, és a központi és mediális AC magokhoz vezetnek . Ez a dopaminerg kapcsolat releváns, mivel kifejezetten a központi mag oldalirányú felosztását célozza meg, egy olyan régiót, amely efferens vetületeket küld a központi mag mediális felosztására, ami viszont visszatér a periaqueductalis szürke anyaghoz, amely potenciálisan veszélyes ingernek kitett állatok “fagyasztó” viselkedését kezeli . A dorsalis raphe magban vannak olyan TH + sejtek is , amelyek a központi AC magba vetülnek, de ezeknek a sejteknek a DAT-tartalmát még nem határozták meg.
a DAT ultrastrukturális lokalizációja a főemlős AC-ben jelenleg nem ismert. Az agykéregben a DAT‐jelölt profilok többsége vékony, nem myelinizált axonoknak felel meg, amelyek ritkán képeznek szinapszist, míg a TH‐jelölt profilok átmérője nagyobb, és a TH‐ir varicositások bőséges vezikulákat tartalmaznak, és gyakran szinapszist képeznek . Következésképpen, Lewis et al. hidd el, hogy a DAT valószínűleg az intervaricose szegmensekre korlátozódik . A dopaminerg rostok specifikus posztszinaptikus célpontjai, amelyek elérik az emberi AC-t, nem ismertek. Számos rágcsálókon végzett tanulmány kimutatta, hogy ezek a rostok szinapszist hoznak létre mind a vetítő neuronokkal, mind az interneuronokkal . Bár a dopaminerg szinapszisok többségét a projekciós neuronok kapják , a CR+ és PV+ interneuron részhalmazokat is ezek a rostok, különösen a PV-t tartalmazó rostok innerválják . A CR+ interneuronok a dopaminerg szinapszisoknak csak 6% – át kapják, míg a PV + sejtek 40% – ot kapnak . A központi és bazális magokban, valamint a paracapsularis interkalált csoportokban a dopaminerg terminálok szimmetrikus szinapszisokat képeznek gyakrabban, mint az aszimmetrikusak .
az AC-ben lévő dopaminerg rostok perineuronális hálókat képeznek a vetítő neuronok és a PV+ interneuronok szóma körül, és ezeknek a hálóknak a PV+ interneuronokkal való érintkezésének 72% – a szinaptikus . Ezek a hálók az összes PV+ interneuron mintegy 10-15% – ában bőségesek, és úgy tűnik, hogy elkerülik a többi interneuron részhalmazt. Ezek a hálók funkcionálisan kapcsolódnak a bazolaterális csoport vetítő neuronjainak aktivitásában megfigyelt erős gátláshoz a dopamin felszabadulása után . Az AC különböző interneuron populációinak dopaminerg beidegzése hozzájárulhat a kondicionált félelem megszerzésében részt vevő hosszú távú potencírozási mechanizmusok indukciójához, amely megköveteli a vetítési neuronok GABAerg interneuron gátlásának elnyomását . A dopamin gátolja a” gyors tüzelésű ” interneuronokat, amelyek egybeesnek a PV+ interneuronokkal , és csökkenti a vetítési neuronok gátlását az oldalsó amygdaloid magban. Újabban, Chu et al. kimutatták, hogy a dopamin blokkolja a GABA felszabadulását a PV+ interneuronokból a D2 típusú preszinaptikus receptorokra ható vetítő neuronokba, de nem befolyásolja a GABA felszabadulását más interneurontípusokba ebből az interneuron populációból . Mind a D1, mind a D2 receptorok blokkolása a bazolaterális csoportban megakadályozza a félelem kondicionált megszerzését .