het humane Amygdaloidcomplex: cellulaire architectuur en dopaminerge innervatie
de dopaminerge innervatie van het humane AC
AC ontvangt een substantiële dopaminerge innervatie die hoofdzakelijk van de ventrale mesencephalic groepen A8, A9 en A10 afkomstig is en dopamine is een belangrijke neurotransmitter in AC die de ingang van informatie door de basolaterale groep moduleert. Bovendien is deze dopaminerge innervatie vereist voor het verwerven, consolideren en uitsterven van angstherinneringen en voor het genereren van geschikte affectieve reacties en, zoals eerder vermeld, zijn disfuncties van dit Dopaminerge systeem voorgesteld als pathogene mechanismen in psychiatrische ziekten zoals schizofrenie en stressgerelateerde aandoeningen . Nauwkeurige kwantitatieve gegevens met betrekking tot de hoeveelheid dopaminerge axonen en hun verdeling in de AC van menselijke donoren die niet waren gediagnosticeerd met neurologische of psychiatrische ziekten vóór hun dood werden verzameld door García‐Amado en Prensa met behulp van dat immunoreactiviteit als marker voor de dopaminerge vezels en stereologische benaderingen. Aangezien de intrinsieke dopamine instabiliteit zijn immunodetectie in hersenenweefsel verhindert dat niet snel door perfusie na de dood van de donor is gefixeerd, hadden vorige studies die bij het analyseren van dopaminerge innervatie van de menselijke AC waren geconcentreerd de TH proteã ne gebruikt om dopaminerge profielen te ontdekken. Nochtans, labelt het eiwit ook noradrenergic en adrenergic vezels in de AC . Aangezien de AC bestaat uit verschillende nucleaire groepen met een breed scala van verbindingen met de cerebrale cortex, hippocampale vorming, basale ganglia, thalamus, hypothalamus, en hersenstam (voor recensie zie Refs. ), is informatie over het gehalte aan dopaminerge axonen in elk van de nucleaire groepen nodig om de interne functionele organisatie van dit complex beter te begrijpen.
het menselijk AC is het doelwit van wijdverbreide dat‐positieve vezels, die dicht en ongelijk verdeeld zijn in elke onderverdeling van dit nucleaire complex (figuren 2 en 3). Voorts heeft hun studie nauwkeurige informatie betreffende de hoeveelheid dat‐ir vezels per neuron in elk amygdaloid Territorium opgeleverd. Zoals door deze auteurs is aangetoond, varieert de hoeveelheid dat‐ir axonen in de menselijke wisselstroom tussen de verschillende kernen van de wisselstroom en varieert ook aanzienlijk in de verschillende onderverdelingen van een gegeven WISSELSTROOMKERN (Figuur 2), hetgeen wijst op functionele variaties tussen deze gebieden.
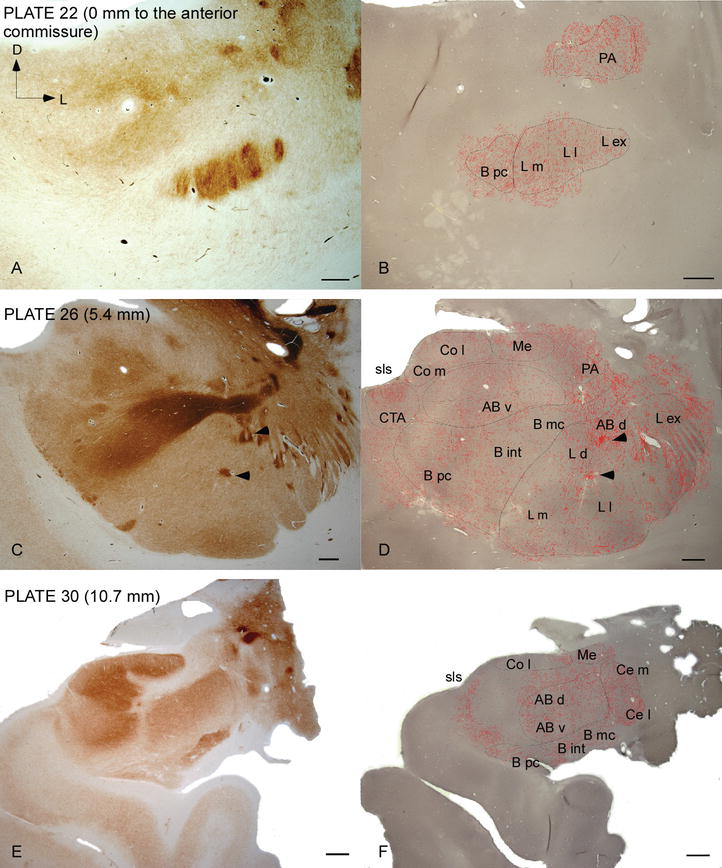
Figuur 2.
verdeling van dat-positieve vezels in het humane AC. Reeks van twee aangrenzende coronale secties gekleurd voor acetylcholinesterase (AChE) (A, C, E) en DAT (B, D, F) op drie anteroposterior niveaus van de AC, met de overeenkomstige platen van Ref. . De stippeling in B, D en F vertegenwoordigt de dat‐positieve axonen getekend met camera lucida op 20× en bovenop dezelfde micrografen gekleurd voor DAT. Pijlpunten in C en D wijzen op vlekken met respectievelijk AChE of met dat verrijkte kleuring. Zie Tabel 1 voor afkortingen. Schaal bar: 1 mm.
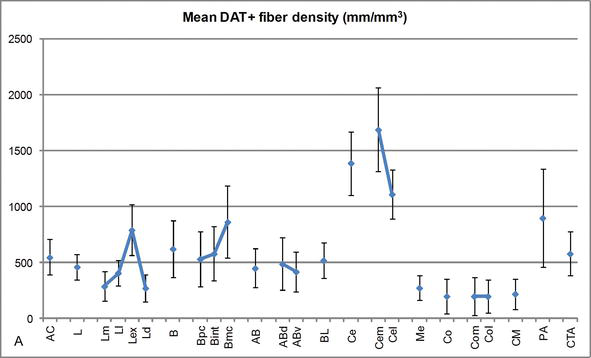
Figuur 3.
Lengtedichtheid van dat-positieve vezels in het menselijke AC. Gemiddelde dat-positieve vezellengte dichtheid voor elke nucleaire groep, kern en nucleaire onderverdeling van de AC. De foutbalken staan voor standaarddeviatie. Zie Tabel 1 voor afkortingen. Gewijzigd van García-Amado en Prensa .
een van de meest opvallende gradiënten in de hoeveelheid dat‐ir‐vezels vond plaats langs de mediolaterale as van de laterale kern: de totale lengte van dat-ir-axonen varieert van bijna 300 mm/mm3 in zijn mediale onderverdeling tot bijna 800 mm/mm3 bij zijn externe (meest laterale) onderverdeling (figuren 2 en 3). Deze grote variatie in de hoeveelheid dat‐ir vezels tussen de mediale en laterale sectoren van de laterale kern kan worden gerelateerd aan hun gedifferentieerde extrinsieke en intrinsieke verbindingen. Zo zou de laterale kern het belangrijkste doel van sensorische informatie uit de externe wereld zijn, en het stuurt zware projecties naar de andere amygdaloã de kernen . De externe onderverdeling van de laterale kern ontvangt de meeste van deze sensorische projecties (figuren 2 en 3), en de informatie stroomt naar de mediale kant van de kern ; bovendien heeft dit AC-gebied de kortste latentie van geconditioneerde reacties opgewekt door sensorische stimuli geassocieerd met ongewenste gebeurtenissen in emotionele leertaken . Aan de andere kant, ontvangt de mediale onderverdeling van de laterale kern informatie van hogere‐orde corticale verwerkingsgebieden . In de hippocampus waren DAT-positieve axonen slechts aanwezig in de buitenste twee derde van de moleculaire laag van de dentaat gyrus, waar de perforantweg eindigt , die erop wijst dat dopamine krachtig en selectief de input van de entorhinale cortex en dus de vroege stadia van hippocampusverwerking kan regelen, zoals het geval zou kunnen zijn voor de sensorische informatie die de AC laterale kern binnendringt.
de centrale kern ontvangt informatie van de overige AC-kernen en is een van de belangrijkste uitgangskernen van de AC . Afdalende projecties van de centrale kern eindigen in een brede mediolaterale regio van mesencephalic dopamine cellen . Op zijn beurt ontvangt deze kern de zwaarste dat‐positieve dopaminerge innervatie van alle AC‐kernen, maar zijn innervatie is niet gelijkmatig verdeeld en neemt duidelijk af langs een mediolaterale gradiënt, een bevinding die overeenkomt met het distributiepatroon van th-ir-vezels . In de basale en bijkomende basale kernen van de AC neemt het gehalte aan dat‐ir vezels af van dorsale naar ventrale sectoren, hoewel deze gradiënt in de laatste veel minder duidelijk is dan in de voormalige kern (zie figuur 2A en B) .
de regulering van de extracellulaire dopaminespiegels wordt gecontroleerd door verschillende mechanismen in verschillende hersengebieden en is waarschijnlijk gerelateerd aan het dat-gehalte. Dus, terwijl het dorsale striatum en de nucleus accumbens een “opname‐gedomineerde” Regulatie vertonen (d.w.z. een waarin dopamine snel wordt teruggewonnen uit de extracellulaire ruimte om zijn actie te beëindigen), vertonen de mediale prefrontale cortex en de AC een “release‐dominated” Regulatie (d.w.z. dopamine wordt langer in de extracellulaire ruimte gehandhaafd) ; deze bevindingen zijn het eens met de observatie dat er meer dat in het striatum is dan in de andere twee structuren .
AC is een belangrijk doelwit voor mesencephalonprojecties die bestaan uit cellen van de substantia nigra pars compacta (dopaminerge A9-groep), het ventrale tegmentale gebied (dopaminerge A10-groep) en het retrorubrale veld (dopaminerge A8-groep) . In het menselijke mesencephalon, dat overvloedig in neuronen gelegen in het laterale ventrale tegmentale gebied en in de substantia nigra pars compacta en is grotendeels afwezig in het mediale ventrale tegmentale gebied . Dat mRNA is overvloediger in de A9 ventrale tier dan in de dorsale tier . De menselijke AC kernen die de meeste dat-ir vezels bevatten komen overeen met die die sterke projecties van het ventrale mesencephalon ontvangen, zoals ook waargenomen bij primaten . Er zijn, niettemin, andere AC gebieden die een hoge dichtheid van dat‐positieve vezels, zoals de laterale onderverdeling van de centrale kern tonen, die innervation van om het even welk deel van de ventrale middenhersenen schijnen niet te ontvangen . Er zijn andere mogelijke bronnen van AC dopamine die buiten de ventrale middenhersenen liggen, maar of zij aan de dat‐ir vezels bijdragen die in AC worden ontmoet of niet, is nog niet duidelijk. De parabrachiale kern projecteert naar de centrale en mediale kernen van het AC en het bevat vermoedelijk dopaminerge neuronen die dat niet dragen . Bovendien missen de neuronen van de parabrachiale kern die projecteren naar de AC ook tyrosinehydroxylase (TH) . De periaqueductale grijze substantie is een andere bron van input aan AC en het bevat dopaminerge neuronen (d.w.z. A11-groep) die dat en project naar de centrale en mediale AC kernen bevatten . Deze dopaminerge verbinding is relevant omdat het specifiek gericht is op de laterale onderverdeling van de centrale kern, een gebied dat efferente projecties stuurt naar de mediale onderverdeling van de centrale kern, die op zijn beurt weer projecteert naar de periaqueductale grijze substantie die “bevriezing” gedrag aanpakt bij dieren blootgesteld aan een potentieel gevaarlijke stimulus . Er zijn ook TH + cellen in de dorsale raphe kern die projecteren naar de centrale AC kern, maar het dat gehalte van deze cellen is nog niet bepaald.
de ultrastructurele lokalisatie van dat in de primaat AC is momenteel onbekend. In de hersenschors, de meeste van de dat‐gelabelde profielen corresponderen met dunne unmyelinated axonen die zelden synapsen vormen, terwijl TH‐gelabelde profielen meer variëren in hun diameter en th‐ir varicosities bevatten overvloedige blaasjes en vormen vaak synapsen . Bijgevolg, Lewis et al. geloven dat DAT waarschijnlijk beperkt is tot de intervaricose segmenten . De specifieke postsynaptische doelen van de dopaminerge vezels die de menselijke AC bereiken zijn niet bekend. Verschillende studies bij knaagdieren hebben aangetoond dat deze vezels synapsen maken met zowel projectie neuronen als interneuronen . Hoewel projectieneuronen de meerderheid van dopaminerge synapsen ontvangen, worden de CR+ en PV+ interneuron subsets ook geïnnerveerd door deze vezels, vooral degenen die PV bevatten . De cr + – interneuronen ontvangen slechts 6% van de dopaminerge synapsen, terwijl de PV+ – cellen 40% ontvangen . In de centrale en basale kernen, evenals in de paracapsulaire intercalatiegroepen, vormen de dopaminerge terminals vaker symmetrische synapsen dan asymmetrische .
dopaminerge vezels in de AC-vorm perineuronale netten rond het soma van de projectieneuronen en de PV+ interneuronen, en 72% van de contacten die deze netten leggen met de PV+ interneuronen zijn synaptisch . Deze netten zijn overvloedig aanwezig in ongeveer 10-15% van alle PV+ interneuronen en lijken andere interneuron-subsets te vermijden. Deze netten zijn functioneel gerelateerd aan de sterke remming waargenomen in de activiteit van de projectie neuronen van de basolaterale groep na dopamine release . De dopaminerge innervatie van de verschillende interneuronpopulaties van de AC zou kunnen bijdragen tot de inductie van langdurige potentiatiemechanismen die betrokken zijn bij geconditioneerde angstverwerving, hetgeen onderdrukking van Gabaerge interneuronremming van projectieneuronen vereist . Dopamine remt de” snelvurende ” interneuronen, die samenvallen met de PV + interneuronen , en vermindert de remming van projectieneuronen in de laterale amygdaloïde kern. Meer recent, Chu et al. hebben aangetoond dat dopamine de GABA-afgifte van PV+ interneuron blokkeert naar projectieneuronen die een werking hebben op type D2 presynaptische receptoren, maar het heeft geen invloed op de afgifte van GABA aan andere interneurontypen uit deze interneuronpopulatie . De blokkade van zowel D1 als D2 receptoren in de basolaterale groep voorkomt angst geconditioneerde acquisitie .